- Research
- Open access
- Published:
Identification key to the Anopheles mosquitoes of South America (Diptera: Culicidae). III. Male genitalia
Parasites & Vectors volume 13, Article number: 542 (2020)
Abstract
Background
Accurate identification of the species of Anopheles Meigen, 1818 requires careful examination of all life stages. However, morphological characters, especially those of the females and fourth-instar larvae, show some degree of polymorphism and overlap among members of species complexes, and sometimes even within progenies. Characters of the male genitalia are structural and allow accurate identification of the majority of species, excluding only those in the Albitarsis Complex. In this key, based on the morphology of the male genitalia, traditionally used important characters are exploited together with additional characters that allow robust identification of male Anopheles mosquitoes in South America.
Methods
Morphological characters of the male genitalia of South American species of the genus Anopheles were examined and employed to construct a comprehensive, illustrated identification key. For those species for which specimens were not available, illustrations were based on published illustrations. Photographs of key characters of the genitalia were obtained using a digital Canon Eos T3i attached to a light Diaplan Leitz microscope. The program Helicon Focus was used to build single in-focus images by stacking multiple images of the same structure.
Results
An illustrated key to South American species of Anopheles based on the morphology of the male genitalia is presented, together with a glossary of morphological terms. The male genitalia of type-specimens of previously poorly documented species were also examined and included in the key, e.g. Anopheles (Anopheles) tibiamaculatus (Neiva, 1906) which has a unique quadrangular-shaped aedeagus with an apical opening.
Conclusions
Male genitalia of South American species of Anopheles possess robust characters that can be exploited for accurate species identification. Distortion that can occur during the dissection and mounting process can obstruct accurate identification; this is most evident with inadvertent damage or destruction of unique features and interferes with correctly assigning shapes of the features of the ventral claspette. In some species, the shape, and anatomical details of the aedeagus also need to be examined for species identification. For members of the Myzorhynchella Series, both ventral and dorsal claspettes possess multiple characteristics that are herein used as reliable characters for species identification.

Background
General introductory comments, distributions and species authors and publication dates are given in Part I [1] of this series of four articles. Keys to the fourth-instar larvae and adult females are provided in Parts II [2] and IV [3], respectively. A list of species treated here is included in Part I [1].
Methods
The primary types (holotypes and paratypes) and other field-collected specimens deposited in the Coleção Entomológica de Referência, Faculdade de Saúde Pública, Universidade de São Paulo, São Paulo, Brazil (FSP-USP), Museo de Entomología, Universidad del Valle, Santiago de Cali, Colombia (MUSENUV) and the US National Mosquito Collection, Smithsonian Institution, Washington, DC, USA (USNMC) were examined to discover characters to be used in the male genitalia key. For species that we could not access, illustrations were based on published illustrations. Photomicrographs of relevant characters for the male genitalia were taken using a digital Canon Eos T3i (Canon, USA), attached to a Diaplan Leitz microscope, using the program Helicon Focus software (https://www.heliconsoft.com/heliconsoft-products/helicon-focus/), which was used to build single in-focus images by stacking multiple images of the same structure. Photomicrographs were further processed in Adobe Photoshop (https://www.photoshop.com/en) to embed names and labels. The institutional sources of specimens are recorded on each photograph. The nomenclature adopted is that of Harbach & Knight [4, 5]. The water marks embedded in photomicrographs of the male genitalia show the institution where the vouchers are deposited, Universidade de Sao Paulo (USP) and Universidad del Valle.
The key includes most of the species registered in South America, except for 15 species that are poorly known. They are the following: Anopheles (Ano.) annulipalpis Lynch Arribálzaga; An. (Ano.) bustamantei Galvão; An. (Ano.) evandroi da Costa Lima; An. (Ano.) pseudomaculipes (Chagas); An. (Ano.) pseudopunctipennis levicastilloi Leví-Castillo; An. (Ano.) pseudopunctipennis neghmei Mann; An. (Ano.) pseudopunctipennis noei Mann; An. (Ano.) pseudopunctipennis patersoni Alvarado & Heredia; An. (Ano.) pseudopunctipennis rivadeneirai Leví-Castillo; An. (Ano.) rachoui Galvão; An. (Ker.) auyantepuiensis Harbach & Navarro; An. (Ker.) boliviensis (Theobald); An. (Ker.) rollai Cova García, Pulido F. & Escalante de Ugueto; An. (Nys.) nigritarsis (Chagas); and An. (Nys.) sanctielii Senevet & Abonnenc. For these species, it will be necessary to conduct field collections in the type-localities and further taxonomic investigations.
Results and discussion
Glossary of morphological terms
The terminology of the male genitalia used in this key follows that of Harbach & Knight [4, 5]. Also known as the male terminalia, Harbach & Knight [4] recommended instead to use “the genitalia” to avoid confusion with other terminal structures. The composite male genitalia are structures formed from elements of the posterior segments IX and X of the abdomen. These modified structures are involved in mating, copulation, and insemination. After emergence of the adult male, the posterior part of the abdomen beyond segment VII makes a 180° rotation. Thus, the ventral segmental surfaces become dorsal in relation to the rest of the abdomen, and vice versa. In Culicidae Meigen, 1818, the male genitalia are therefore inverted in relation to the female genitalia. This means that when coupling occurs, both individuals have the same upright orientation, instead of the male ending up vulnerable, and upside down in relation to the female, as is the case in the family Tabanidae Latreille, 1802 (horse flies), which do not have male genital rotation [6]. This phenomenon must be considered when describing the position of the various elements of the genitalia.
Tergum IX, which usually varies little, can exhibit useful morphological variation in certain species. Species of the Arribalzagia Series of the subgenus Anopheles Meigen, 1818 possess ninth tergal lobes (IX-Te lobes) of variable size and development, features which can be useful for species recognition. The internal margin of tergum IX is attached to the proctiger that is formed by tergum X, the cerci, the cercal sclerites and the paraprocts. Dorsally there are two sclerotized plates called the cercal sclerites. The two structures attached laterally on the most posterior part of the abdomen are called the gonocoxopodites. They are adapted to facilitate insemination by grasping the female during copulation. The gonocoxopodites are composed of a proximal gonocoxite and a distal gonostylus.
The gonocoxite (Fig. 1) is a large, relatively long, and somewhat conical structure, much wider than the gonostylus. The external ventral surface is convex, while the internal surface is slightly concave, especially basally at the attachment of the claspettes [7]. The gonocoxites bear a large number of scales and setae, the larger of which are sometimes called spines; for purposes of this key, the term seta will be used. One or two parabasal setae are inserted on the dorsobasal portion of the gonocoxite. In species of the subgenera Nyssorhynchus Blanchard, 1902 and Kerteszia Theobald, 1905, the setae are inserted on prominent parabasal lobes that are situated dorsobasally (in a prerotational sense). The parabasal lobes are absent in Neotropical species belonging to the subgenera Anopheles, Lophopodomyia Antunes, 1937 and Stethomyia Theobald, 1902. The parabasal setae are instead inserted directly on the surface or on relatively small projections on the surface of the gonocoxite. In species of the subgenus Stethomyia, the parabasal setae are absent.

Schematic drawing of the male genitalia of An. pseudopunctipennis Theobald, 1901. The proctiger (anal lobe) was removed to permit an unobstructed view of the ventral structures (redrawn after Komp [7])
Species of the subgenera Nyssorhynchus (Fig. 2) and Kerteszia have a single dorsally directed seta that is inserted on the parabasal lobe. The Neotropical species of the subgenera Anopheles and Lophopodomyia have two parabasal setae (Fig. 3b, c). The internal seta is inserted on the ventral surface of the gonocoxite, on the distal half or near mid-length. Species of the subgenera Nyssorhynchus and Kerteszia have a pair of accessory setae inserted on the dorsal surface of the gonocoxite (Figs. 2, 3a).

Dorsal view of the male genitalia of An. albimanus Wiedemann. Segment IX and the proctiger (anal lobe) were removed to facilitate the observation of structures that occupy the ventral position

Gonocoxites showing the accessory setae, internal seta and parabasal setae of species of the subgenera Nyssorhynchus (a), Anopheles (b), Stethomyia (c). b and c redrawn after Komp [7]
The gonostylus corresponds to the stylus of the gonocoxopodite. It is a well-sclerotized structure, moveable and articulated, on or near the apex of the gonocoxite. It is somewhat thickened and curved. At its apex is a small spiniform structure called the gonostylar claw [8] (Fig. 3).
Attached to the internal surface of the gonocoxite is the claspette. This is a membranous structure, usually divided into ventral and dorsal lobes, both exhibiting great variability according to subgenus and species within the subgenera (Fig. 4). For purposes of this key, the ventral and dorsal lobes of the claspette [4] are referred to as the ventral claspette and dorsal claspette, respectively. These are terms also used by Faran [9].

Ventral and dorsal lobe claspettes of the male of An. neomaculipalpus Curry, 1931(redrawn after Komp [7])
In species of the subgenera Anopheles, Lophopodomyia and Kerteszia, the dorsal claspette is divided into ventral and dorsal lobes. These lobes exhibit interspecific variability in the form of the setae, as well as the size and distribution of spicules, which makes them useful in taxonomy. In species of the subgenera Anopheles (Fig. 4), Lophopodomyia and Kerteszia, the ventral lobe is divided apically, with the distance between them being more pronounced in species of the subgenera Kerteszia. In species of the subgenus Nyssorhynchus, the ventral claspette is not subdivided, instead the two ventral claspettes are fused and the composite structure occupies a median position between the gonocoxites. The structure is rich in morphological variation and is therefore useful for species identification. In some taxa, the ventral claspette is smooth (i.e. without spicules) while in others the spicules can be short or long and variously distributed on the claspette.
The apex of the ventral claspette can be rounded, truncated or angular, and, in some species can bear apicolateral expansions that resemble lobes. The presence of these apicolateral lobes in An. (Nys.) triannulatus (Neiva & Pinto, 1922) allows separation of this species from the otherwise morphologically similar An. (Nys.) halophylus Silva-do- Nascimento & Lourenço-de-Oliveira, 2002. Other structures that make up parts of the ventral claspette and are employed in taxonomy treatments include the preapical plate and basoventral lobes (Fig. 5). In species of the subgenera Nyssorhynchus, Kerteszia, Anopheles and Lophopodomyia, there is a short basal portion on the dorsal claspette upon which variable numbers of setae are attached dorsoventrally. These setae are variable in form, point of insertion, development, and quantity (Fig. 6). In species of the subgenus Stethomyia, the dorsal claspette is absent and the ventral claspette is columnar, with two subdivisions that support apical setae that are variously developed [7].

Ventral claspette of An. strodei Root, 1926 (redrawn after Faran & Linthicum [10])

Basal portion of dorsal claspette of species of Anopheles (Nyssorhynchus). a An. braziliensis (Chagas, 1907). b An. argyritarsis Robineau-Desvoidy, 1827 (redrawn after Faran & Linthicum [10])
The aedeagus is part of the phallosome, which includes, in addition to the aedeagus, the parameres and the basal pieces. The aedeagus is articulated basally to the parameres, which are connected to the basal pieces by an acetabulum that is on the median lateral area. The basal pieces, responsible for movement of the aedeagus during copulation, are connected to the gonocoxal apodemes. The aedeagus is the central organ of the phallosome and serves as the intromittent organ [4]. In species of Anopheles, the aedeagus is a tubular structure, dorsally curved, with the walls unequally sclerotized and with a circular opening near or at the apex. The apical part of the aedeagus is variable in form and development. In species of the subgenus Nyssorhynchus, the apical part of the aedeagus is variable, and often used in species identification. The presence of leaflets subapically on the aedeagus, as well as the number of these structures, their form, development, and presence of marginal serrations, permit identification of many species of the genus (Fig. 7).

Aedeagus of the male genitalia. a Subgenus Anopheles. b Subgenus Nyssorhynchus (redrawn after Komp [7])
In species of the subgenus Anopheles, the apical leaflets, when present, can vary in number, position, form, and development. Some species possess a single pair of leaflets that can have smooth or serrate margins, be uniformly or unevenly sclerotized, and be short or long. In species of the subgenera Kerteszia and Nyssorhynchus, the leaflets may be present or absent. When present, they occur as a single subapical pair.
The morphological key using the external characters of the male genitalia can aid in identifying species of the genus Anopheles of the South America. Unnamed species of the known complexes can be identified as morphologically similar valid species. In the key, species complex is labelled as (s.l.). For these groups, further investigations will be necessary to define characters of the male genitalia for accurate identification. The key was modified from [10, 11, 13, 14], with additional characters provided herein.
Key for the identification of species of the genus Anopheles of South America based on characters of the male genitalia
-
1a
Gonocoxite without parabasal setae (Fig. 8a)…..2
Fig. 8 
a An. thomasi Shannon, 1933. b An. antunesi Galvao & Franco do Amaral, 1940
-
1b
Gonocoxite with 1 or 2 parabasal setae (Fig. 8b)…..6
-
2a (1a)
Ventral claspette with 2 spiniform setae (Fig. 9a)…..3
Fig. 9 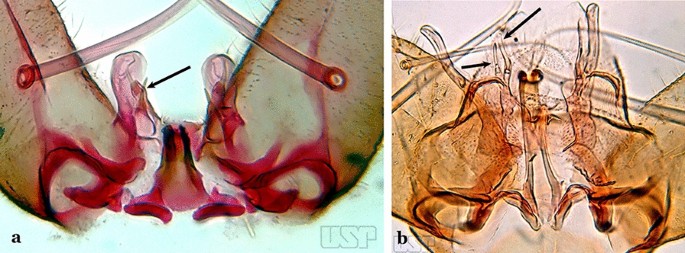
a An. kompi Edwards, 1930. b An. thomasi
-
2b
Ventral claspette with 1 spatulate and 1 spiniform seta (Fig. 9b)…..4
-
3a (2a)
Accessory seta inserted on proximal third of gonocoxite (Fig. 10)….. An. kompi
Fig. 10 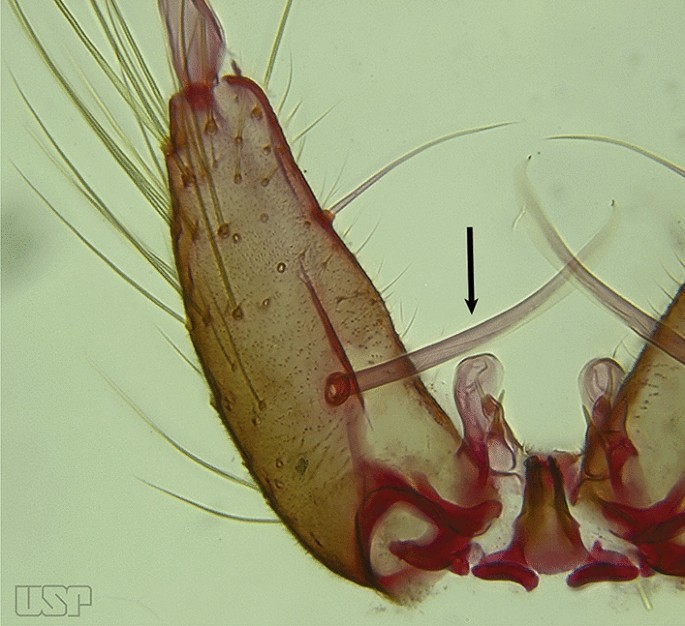
An. kompi
-
3b
Accessory seta inserted near middle of gonocoxite….. An. canorii
-
4a (2b)
Dorsal claspette with longest subdivision lacking a subapical projection in form of a beak (Fig. 11a)…..An. thomasi
Fig. 11 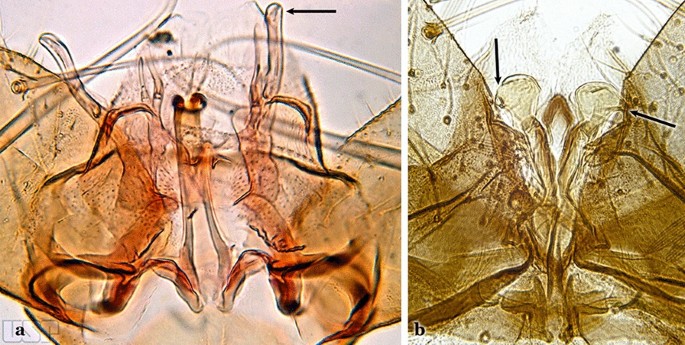
a An. thomasi. b An. nimbus (Theobald, 1902)
-
4b
Dorsal claspette with longest subdivision having a subapical projection in form of a long or short beak (Fig. 11b)…..5
-
5a (4b)
Dorsal claspette with longest subdivision having a subapical projection in form of a long beak (Fig. 12a)….. An. acanthotorynus
Fig. 12 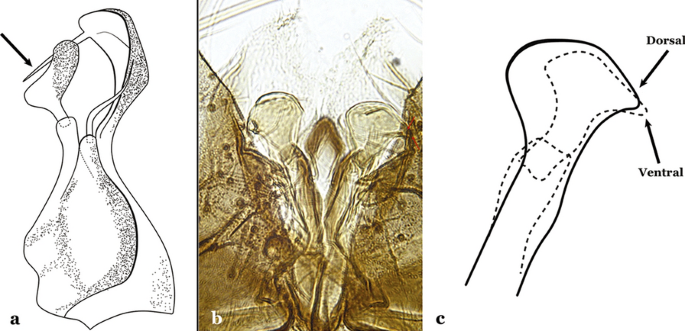
a An. acanthotorynus Komp, 1937 (redrawn after Komp [12]). b, c An. nimbus (c drawn from b)
-
5b
Dorsal claspette with longest subdivision having a subapical projection in form of a short beak (Fig. 12b)…..An. nimbus
-
6a (1b)
Gonocoxite with 2 accessory setae and 1 internal seta (Fig. 13a)…..7
Fig. 13 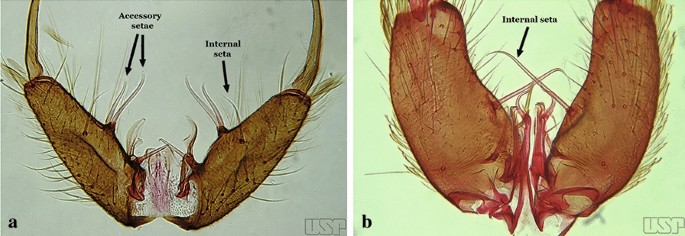
a An. braziliensis. b An. costai da Fonseca & da Silva Ramos, 1940
-
6b
Gonocoxite without accessory setae, internal seta present or absent (Fig. 13b)…..53
-
7a (6a)
Gonocoxite with parabasal seta inserted on a basodorsal lobe (Fig. 14a)…..8
Fig. 14 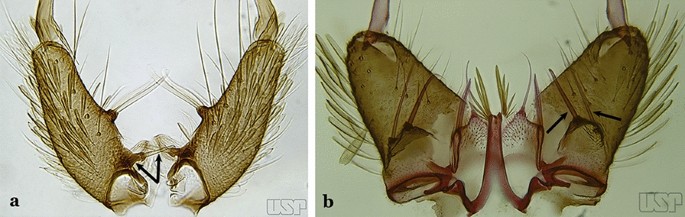
a An. darlingi Root, 1926. b An. peryassui Dyar & Knab, 1908
-
7b
Gonocoxite with parabasal seta inserted directly on the surface of its proximal third (Fig. 14b)…..48
-
8a (7a)
Parabasal seta long and curved, apex truncate; internal seta inserted between accessory and parabasal setae (Fig. 15a); sternum IX with a median longitudinal apodeme (Fig. 15b) (Kerteszia)…..9
Fig. 15 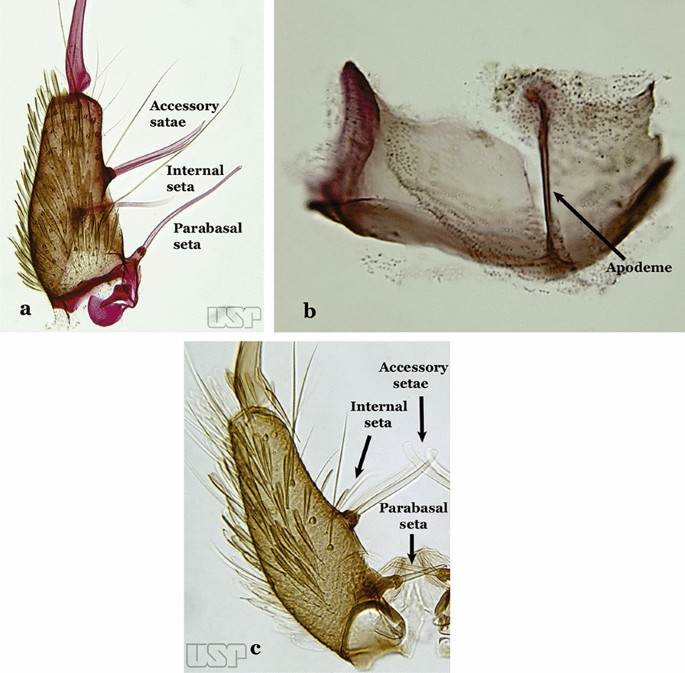
a, b An. homunculus Komp, 1937. c An. darlingi Root, 1926
-
8b
Parabasal seta short and strong, apex ending in a small hook or tapering to apex; accessory setae inserted between internal and parabasal setae (Fig. 15c); sternum IX without a median longitudinal apodeme (Nyssorhynchus)…..17
-
9a (8a)
Aedeagus without leaflets (Fig. 16a)…..10
Fig. 16 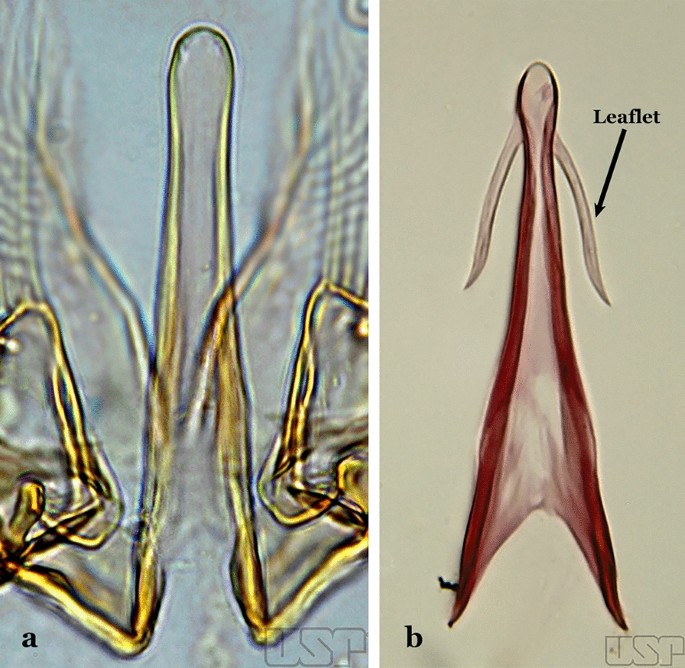
Aedeagus of An. neivai (Lane & Coutinho, 1940) (a) and An. cruzii Dyar & Knab, 1908 (b)
-
9b
Aedeagus with a pair of subapical leaflets (Fig. 16b)…..14
-
10a (9a)
Ventral claspette smooth except for 4–11 strong spicules, and/or less developed spicules along median margin (Fig. 17a)…..An. bambusicolus
Fig. 17 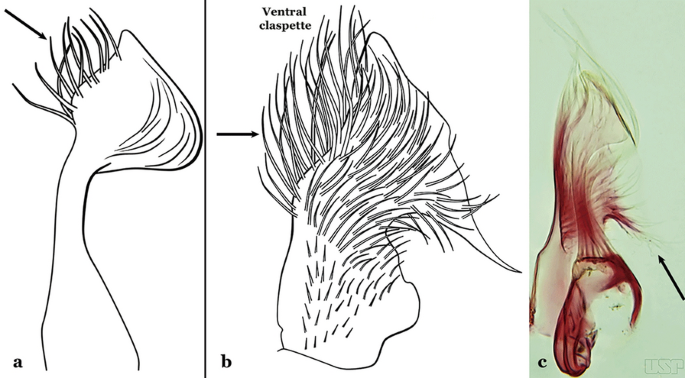
a Ventral claspette of An. bambusicolus Komp, 1937. b, c An. homunculus (b redrawn after Zavortink [13])
-
10b
Ventral claspette moderately or densely spiculose, spicules distributed over its entire surface (Fig. 17b)…..11
-
11a (10b)
Tergum VIII without broad median scales (Fig. 18)…..12
Fig. 18 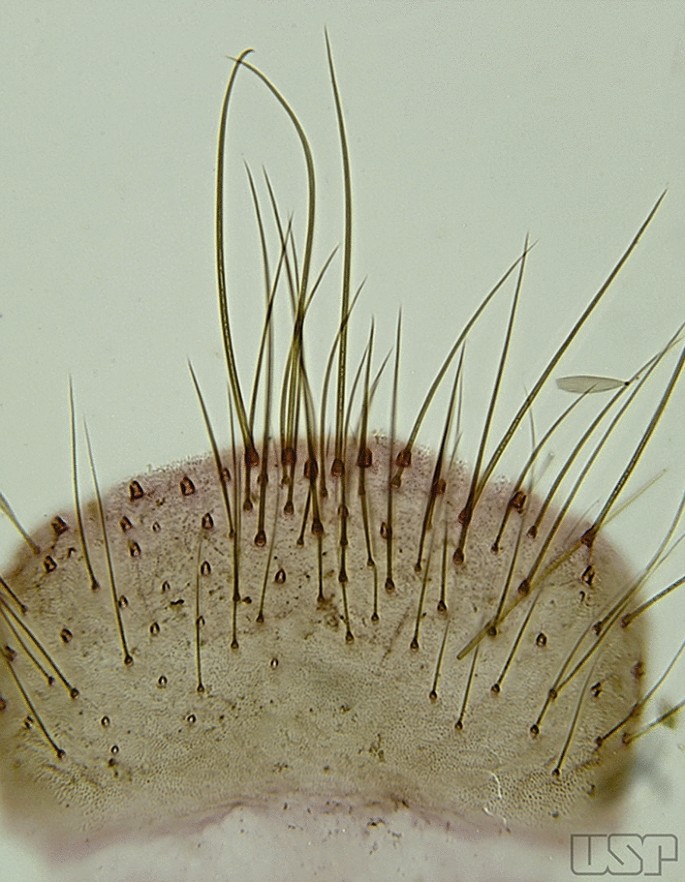
An. homunculus
-
11b
Tergum VIII with numerous broad median scales…..13
-
12a (11a)
Ventral claspette with a straight lateral expansion, not curved ventroposteriorly; gonocoxite with internal seta flattened, wider near apex (Fig. 19a)….. An. neivai (s.l.)
Fig. 19 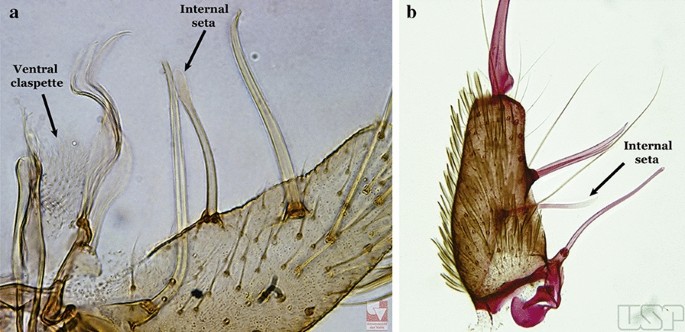
a An. neivai. b An. homunculus
-
12b
Ventral claspette with a large lateral expansion, curved ventroposteriorly, forming a sharp point directed anteriorly (Fig. 17b); internal seta flattened but only slightly wider near apex (Fig. 19b)….. An. homunculus (in part)
-
13a (11)
Ventral claspette with a rounded lateral expansion, not forming an anteriorly directed lobe (Fig. 20a); internal seta flattened apically…..An. pholidotus & An. gonzalezrinconesi
Fig. 20 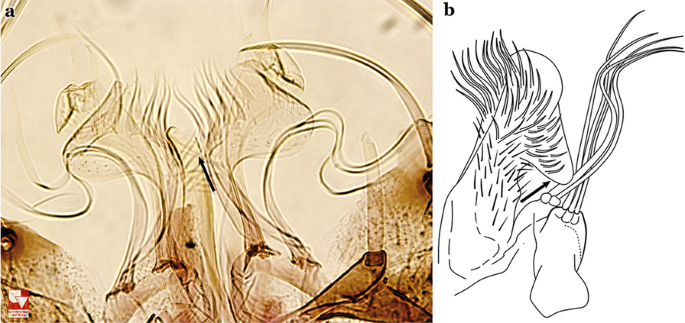
a An. pholidotus Zavortink, 1973. b An. lepidotus Zavortink, 1973 (redrawn after Zavortink [13])
-
13b
Ventral claspette slightly emarginated and amply expanded laterally, forming a rounded anteriorly directed lobe (Fig. 20b); internal seta not flattened apically…..An. lepidotus
-
14a (9b)
Ventral claspette with a large lateral expansion, curved posteroventrally and forming a sharp anteriorly directed point (Fig. 21a)….. An. homunculus (in part)
Fig. 21 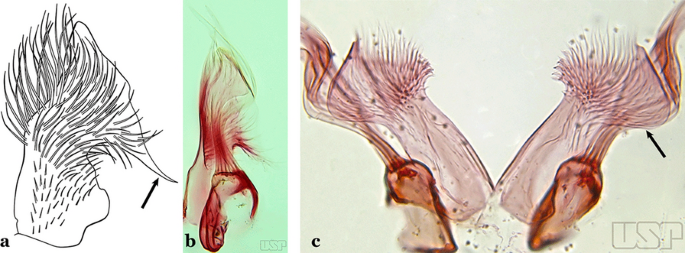
a, b An. homunculus (a redrawn after Zavortink [13]). c An. laneanus Correa & Cerqueira, 1944
-
14b
Ventral claspette with a lateral expansion not forming a sharp anteriorly directed point or with a rounded posteriorly directed lobe (Fig. 21b)…..15
-
15a (14b)
Ventral claspette with a lateral expansion varying from more or less rounded to sinuous on lateral margin, not posteriorly curved (Fig. 22a)….. An. cruzii (s.l.)
Fig. 22 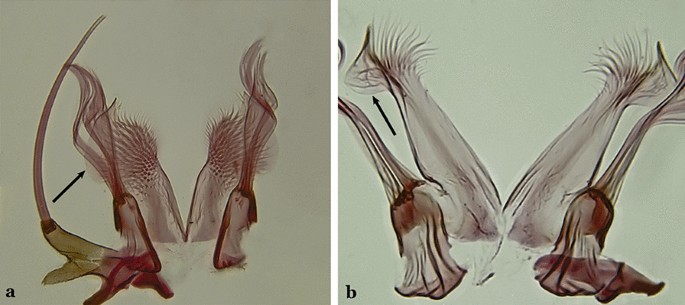
a An. cruzii. b An. bellator Dyar & Knab, 1906
-
15b
Ventral claspette with a rounded lateral expansion, curved posteroventrally, densely spiculate medially (Fig. 22b)…..16
-
16a (15b)
Ventral claspette spiculose medially, with few short spicules laterally; spiculose portion up to 0.25 length of ventral claspette (Fig. 23a)…..An. bellator
Fig. 23 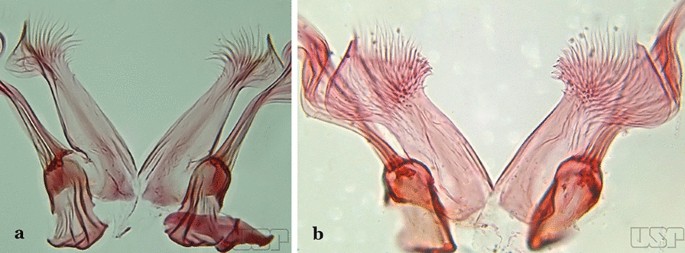
a An. bellator. b An. laneanus
-
16b
Ventral claspette densely spiculose medially, with many short spicules laterally that extend to lateral margin; spiculose portion approximately 0.40 length of ventral claspette (Fig. 23b)…..An. laneanus
-
17a (8b)
Ventral claspette without spicules or setae or with only small spicules mesally on basoventral surface (Fig. 24a)…..18
Fig. 24 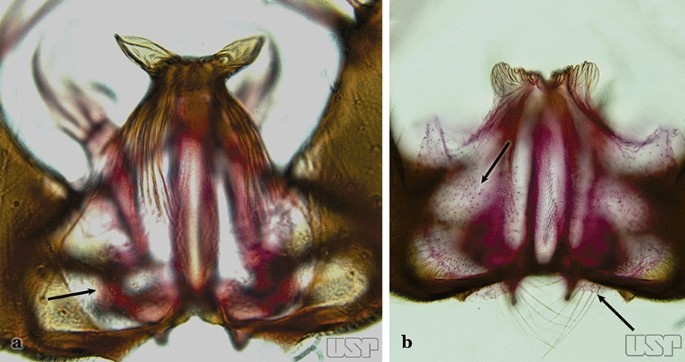
a An. triannulatus. b An. strodei
-
17b
Ventral claspette with spicules or setae at least on basal lobe (Fig. 24b)…..34
-
18a (17a)
Ventral claspette with a laterally expanded apex, forming a well- developed apicolateral ear-like lobe (Fig. 25a)…..19
Fig. 25 
a An. triannulatus. b An. guarani Shannon, 1928
-
18b
Ventral claspette with or without an apicolateral expansion, if expansion present it is never ear-like (Fig. 25b)…..20
-
19a (18a)
Apicolateral lobes moderately narrow basally and directed laterally or posterolaterally (Fig. 26a)….. An. triannulatus
Fig. 26 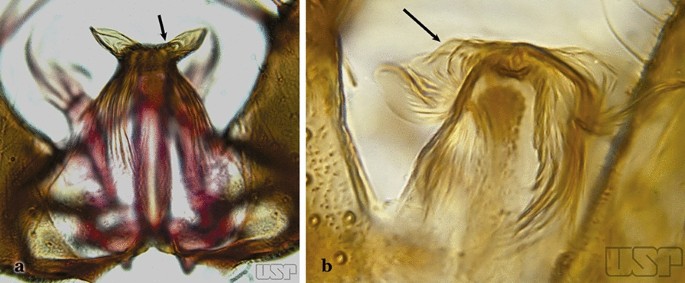
a An. triannulatus. b An. halophylus
-
19b
Apicolateral lobes wide basally and generally directed anteriorly (Fig. 26b)….. An. halophylus
-
20a (18b)
Aedeagus with a pair of well-sclerotized subapical leaflets (Fig. 27a)…..21
Fig. 27 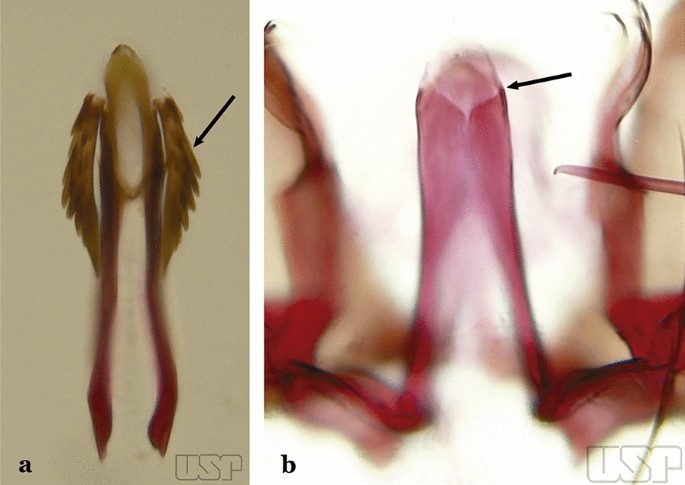
a An. guarani. b An. albitarsis Lynch Arribálzaga, 1938
-
20b
Aedeagus without leaflets (Fig. 27b)…..31
-
21a
Apex of aedeagus sclerotized centrally, hyaline laterally, ending in a dorsally curved hook (Fig. 28a)…..An. parvus
Fig. 28 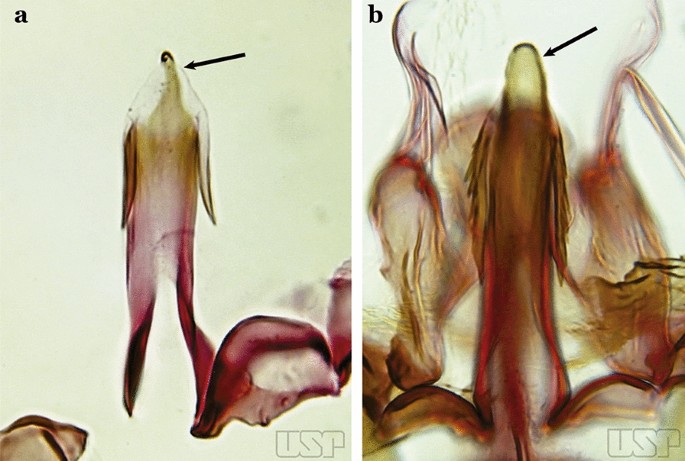
a An. parvus (Chagas, 1907). b An. antunesi Galvão & Franco do Amaral, 1940
-
21b
Apex of aedeagus not as above, straight, without an apical hook (Fig. 28b)…..22
-
22a (21b)
Dorsal claspette with 2 apical and 1 subapical setae (Fig. 29a)…..23
Fig. 29 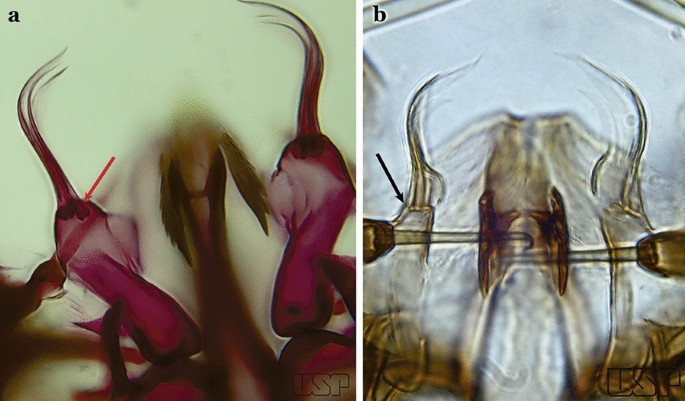
a An. atacamensis González & Sallum, 2010. b An. darlingi
-
22b
Dorsal claspette with all setae inserted apically (Fig. 29b)…..28
-
23a (22a)
Proctiger smooth or with minute spines laterally (Fig. 30a)…..24
Fig. 30 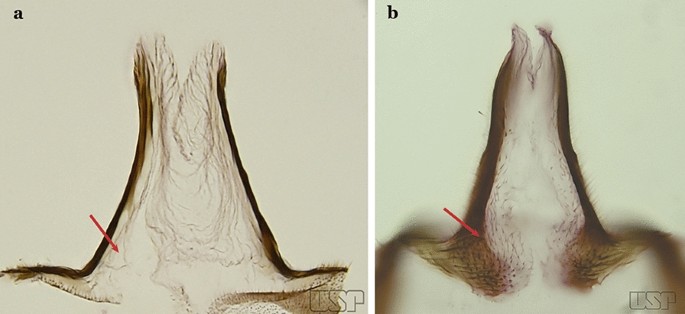
a An. antunesi. b An. atacamensis
-
23b
Proctiger spiculose, at least basally (Fig. 30b)…..25
-
24a (23a)
Ventral claspette with a narrow apex; aedeagus with subapical leaflets positioned parallel to its longitudinal axis (Fig. 31a)…..An. antunesi
Fig. 31 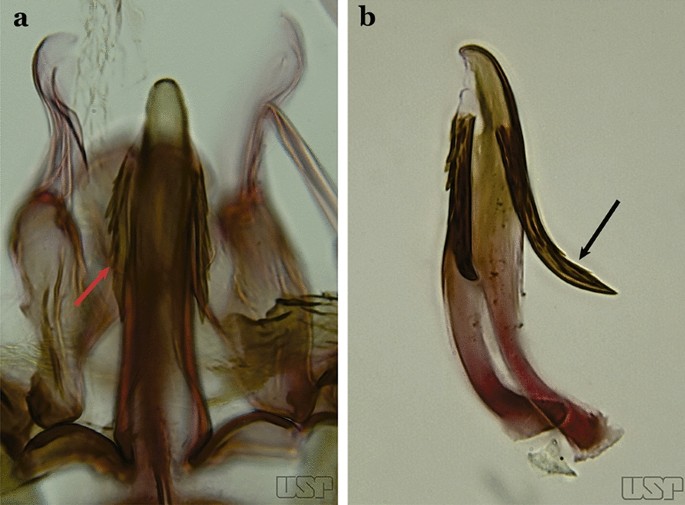
a An. antunesi. b An. pristinus Nagaki & Sallum, 2010
-
24b
Ventral claspette with a rounded apex; aedeagus with subapical leaflets at about a 25° angle in relation to its longitudinal axis (Fig. 31b)….. An. pristinus
-
25a (23b)
Internal seta of gonocoxite straight (Fig. 32a)…..26
Fig. 32 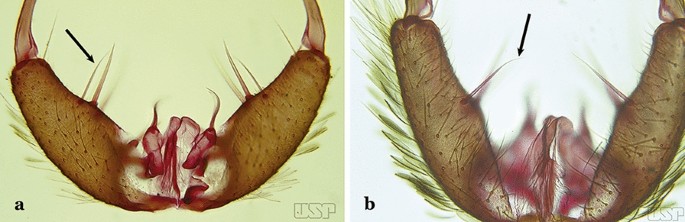
a An. pictipennis (Philippi, 1865). b An. lutzii Cruz, 1901
-
25b
Internal seta of gonocoxite distally curved (Fig. 32b)…..27
-
26a (25a)
Aedeagus with a conical or rounded apex; ventromedian subtriangular projection forming a large deep arch (Fig. 33a)…..An. atacamensis
Fig. 33 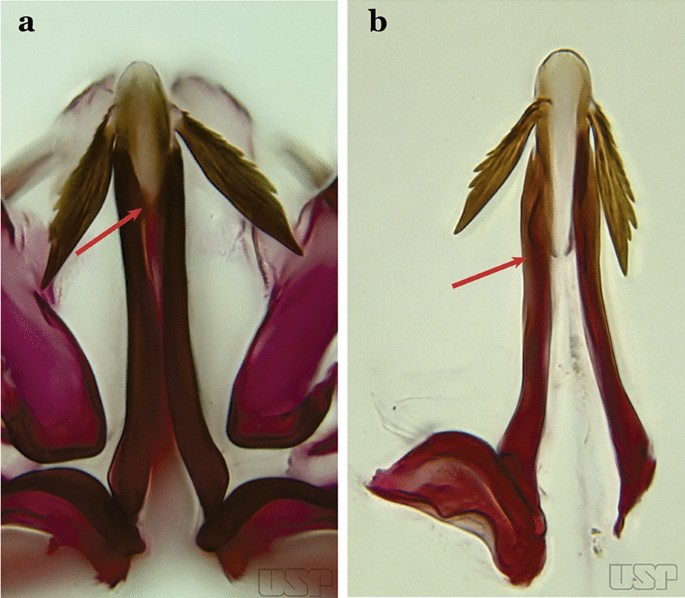
a An. atacamensis. b An. pictipennis
-
26b
Aedeagus with a rounded apex; ventromesal subtriangular projection forming an open narrow arch (Fig. 33b)….. An. pictipennis
-
27a (25b)
Ventral claspette with a large basoventral lobe (Fig. 34a); aedeagus with subapical leaflets directed laterally, forming an angle of about 45° in relation to its longitudinal axis (Fig. 34b)…..An. lutzii
Fig. 34 
a, b An. lutzii. c, d An. guarani Shannon, 1928
-
27b
Ventral claspette with a small basoventral lobe (Fig. 34c); aedeagus with subapical leaflets directed posteriorly, positioned approximately parallel to its longitudinal axis (Fig. 34d)….. An. guarani
-
28a (22b)
Apex of aedeagus longer than wide; ventromesal triangular projection of aedeagus envelops all of subapical region, forming a distinct collar (Fig. 35a); dorsal seta of dorsal claspette with a well-defined basomedian projection (Fig. 29b); apex of ventral claspette moderately wide and truncate; median sulcus small, often indistinguishable (Fig. 35b)….. An. darlingi
Fig. 35 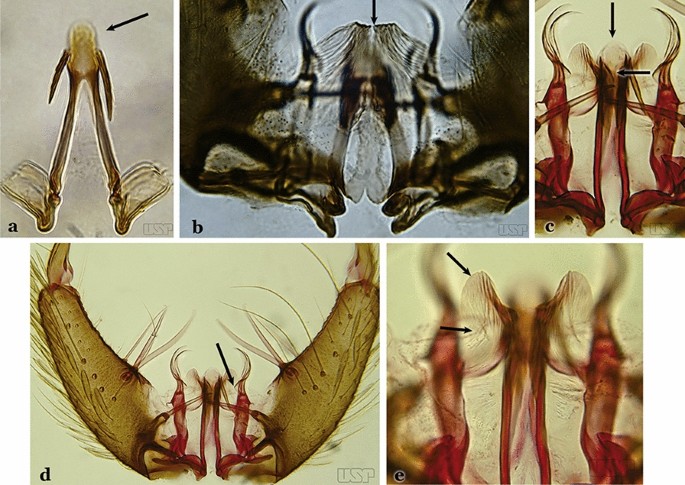
a, b An. darlingi. c, d, e An. lanei Galvão & Franco do Amaral, 1938
-
28b
Apex of aedeagus wider than long (Fig. 35c); ventromesal triangular projection of aedeagus usually absent, when present, it is in form of a collar; dorsal seta of dorsal claspette with or without a well-defined basomedial projection (Fig. 35d); apex of ventral claspette variable; median sulcus large, clearly distinguishable (Fig. 35e)…..29
-
29a (28b)
Ventral claspette with apex laterally expanded into a large rounded lobe, directed posteriorly, distal third distinctly narrowed (Fig. 35e); aedeagus with relatively long slightly serrated subapical leaflets (Fig. 36a); aedeagus without ventromedial triangular projection; dorsal seta of dorsal claspette with a prominent basomedial projection (Fig. 35d)….. An. lanei
Fig. 36 
a An. lanei. b, c An. argyritarsis
-
29b
Ventral claspette with apex not expanded or slightly expanded laterally, distal third not narrowed (Fig. 36b); aedeagus with strongly serrated subapical leaflets (Fig. 35a); aedeagus with a distinct ventromedial triangular projection (Fig. 35a); dorsal seta of dorsal claspette without a basomedial projection, or not prominent if present (Fig. 36c)…..30
-
30a (29b)
Ventral claspette with 2 rounded apicolateral expansions; median sulcus wide, distinct, sclerotized medially (Fig. 37a)…..An. sawyeri
Fig. 37 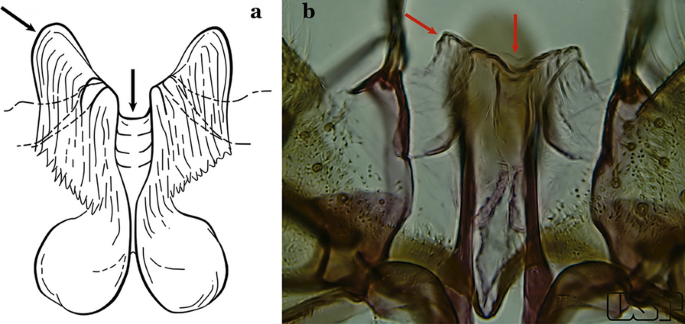
a An. sawyeri Causey, Deane, Deane & Sampaio, 1943. b An. argyritarsis
-
30b
Ventral claspette without apicolateral expansions; median sulcus narrow, indistinct (Fig. 37b)…..An. argyritarsis
-
31a (20b)
Ventral claspette with a pair of sack-like dilations on its ventral surface, situated just below apex; preapical plate small, well sclerotized (Fig. 38a)…..An. albimanus
Fig. 38 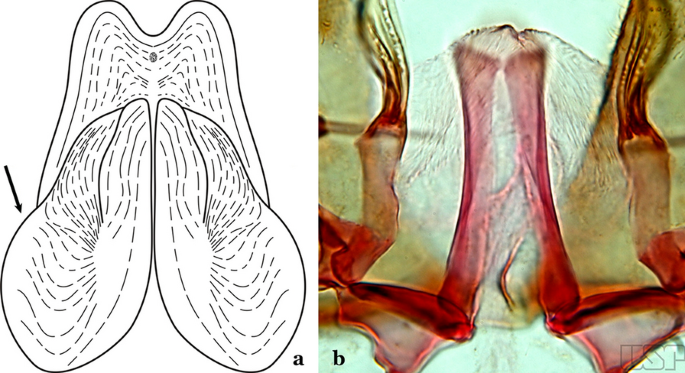
a An. albimanus (redrawn after Faran [9]). b An. marajoara Galvão & Damasceno, 1942
-
31b
Ventral claspette not as above (Fig. 38b)…..32
-
32a (31b)
Ventral claspette with a rounded apex, triangular or conical in appearance (Fig. 39a)…..An. albitarsis, An. deaneorum, An. janconnae, An. marajoara & An. oryzalimnetes
Fig. 39 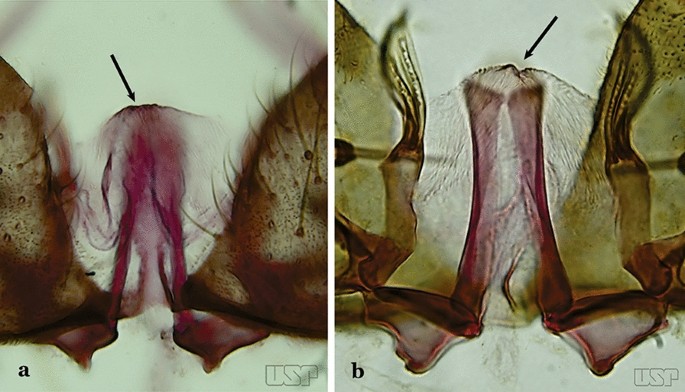
a An. albitarsis Lynch Arribálzaga, 1878. b An. marajoara
-
32b
Ventral claspette with apex truncate or slightly, rounded trapezoidal in appearance (Fig. 39b)…..33
-
33a (32b)
Parabasal seta with a hook-like apex, inserted on a well-developed parabasal lobe (Fig. 40a); dorsal seta of dorsal claspette with basomedial projection well-developed; ventral claspette truncate; preapical plate present, distinct (Fig. 40b)…..An. braziliensis
Fig. 40 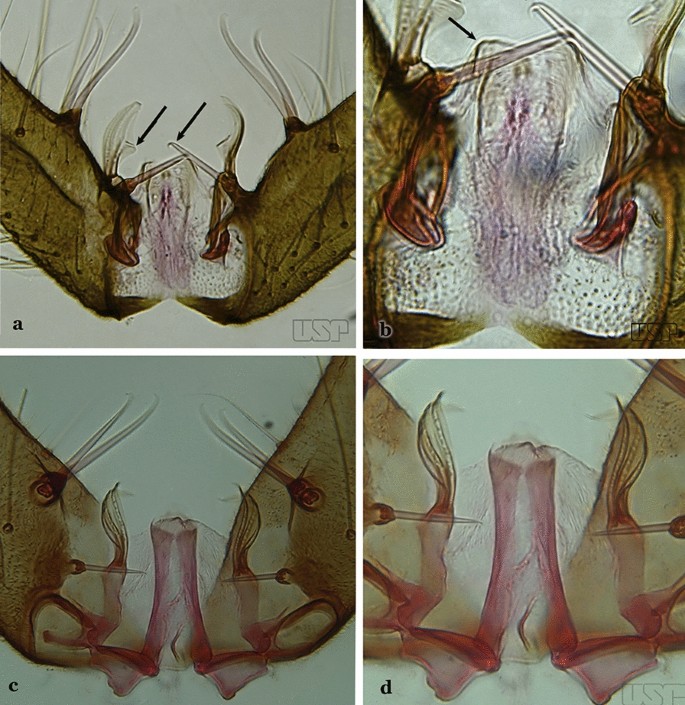
a, b An. braziliensis. c, d An. marajoara
-
33b
Parabasal seta with a pointed apex, curved, inserted on a reduced parabasal lobe (Fig. 40c); dorsal seta of dorsal claspette without basomedial projection; ventral claspette slightly rounded at apex; preapical plate absent (Fig. 40d)…..An. marajoara
-
34a (17b)
Ventral claspette with a smooth apex, expanded, either rugose or distinctly striate (Fig. 41a)…..35
Fig. 41 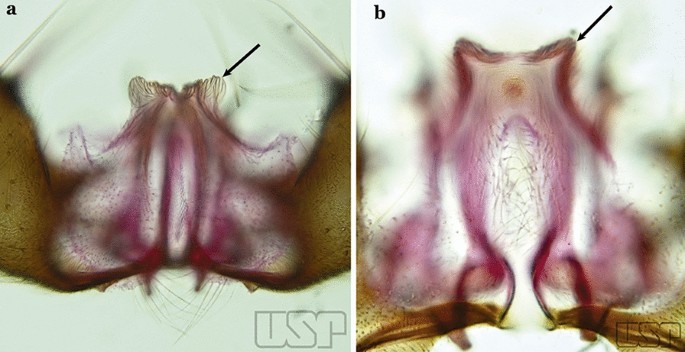
a An. strodei. b An. nuneztovari Gabaldon, 1940
-
34b
Ventral lobe of claspette with a spiculose apex, slightly or not expanded, rugose or striate (Fig. 41b)…..39
-
35a (34a)
Ventral claspette small, apex moderately expanded laterally, apicolateral margins pointed or moderately angular (Fig. 42a); basoventral lobe of ventral claspette curved in a medial direction (Fig. 42b); preapical plate well sclerotized…..An. benarrochi
Fig. 42 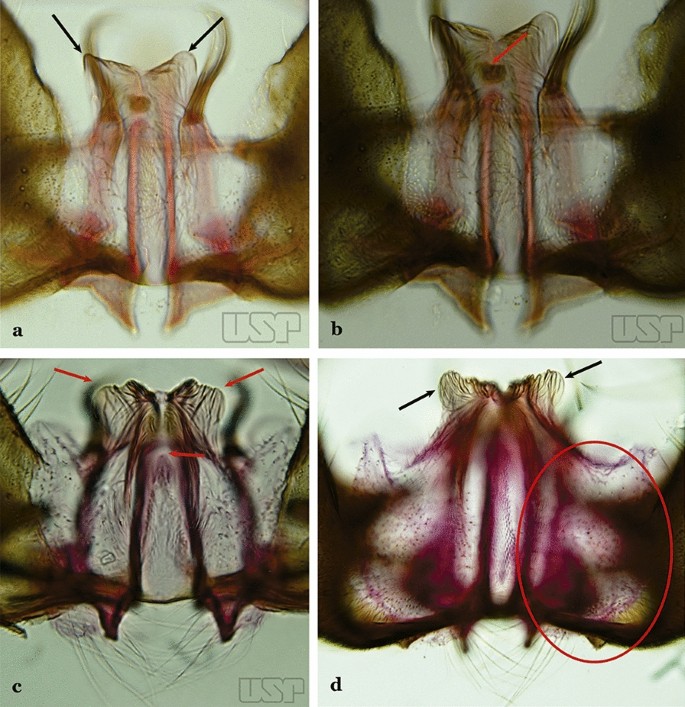
a, b An. benarrochi Gabaldon, Cova-Garcia & Lopez, 1941. c An. rondoni (Neiva & Pinto, 1922). d An. strodei
-
35b
Ventral claspette with apex strongly expanded laterally, apicolateral margins in form of rounded lobes (Fig. 42c, d); basoventral lobe of ventral claspette not as above (Fig. 42d); preapical plate poorly sclerotized or absent (Fig. 42c)…..36
-
36a (35b)
Ventral claspette with expanded apicolateral margins, quadrangular, lateral margins convex and apical margin slightly concave (Fig. 43a); basoventral lobe large, elongate distally (Fig. 43b); preapical plate poorly developed (Fig. 43a)…..An. rondoni
Fig. 43 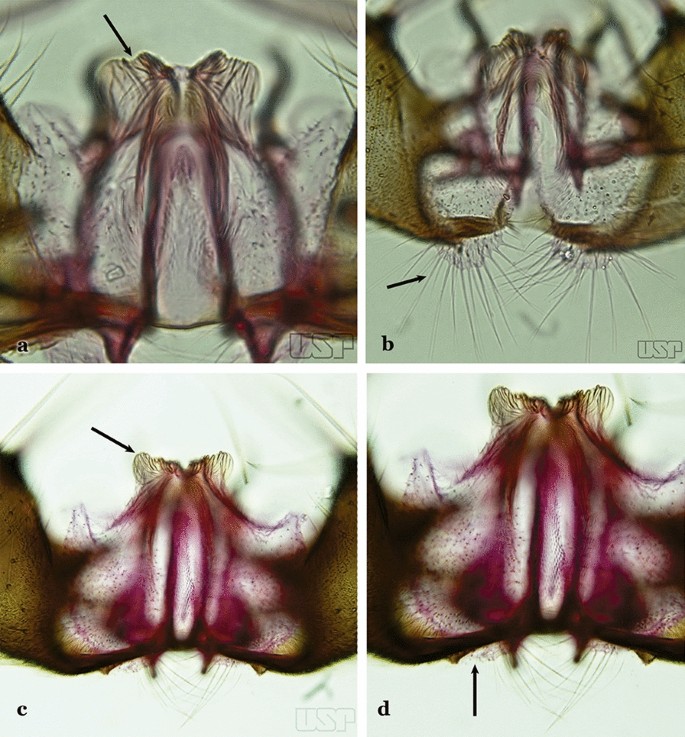
a, b An. rondoni. c, d An. strodei
-
36b
Ventral claspette with apicolateral margins projected as large rounded lobes, lateral margins convex and apical margin slightly concave (Fig. 43c); basoventral lobe of ventral claspette large, apically rounded (Fig. 43d); preapical plate slightly to moderately defined (Fig. 43c)…..37
-
37a (36b)
Ventral claspette with apicolateral margins strongly expanded laterally, forming well-developed lobes; ventral claspette without spicules on dorsal and lateral surfaces of apical half (Fig. 44a)….. An. arthuri (s.l.)
Fig. 44 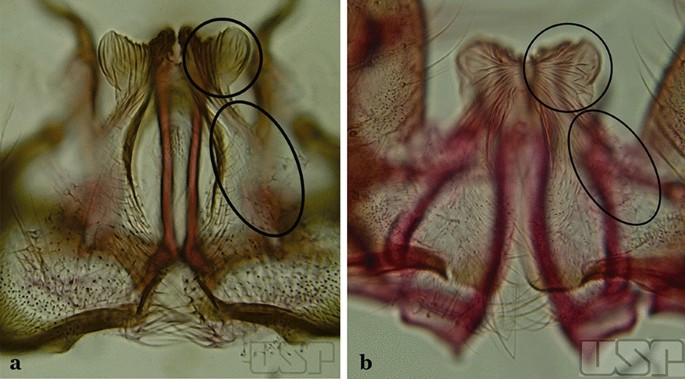
a An. arthuri Unti, 1941. b An. albertoi Unti, 1941
-
37b
Ventral claspette with apicolateral margins moderately expanded laterally, forming moderately developed lobes; ventral claspette with spicules on dorsal and lateral surfaces of apical half (Fig. 44b)…..38
-
38a (37b)
Spicules of lateral and dorsal surfaces of apical half of ventral claspette extend to 0.8 of apical portion (Fig. 45a); basoventral lobe of ventral claspette with long well-developed spicules, all similar in size and development, arranged along distal margin…..An. strodei
Fig. 45 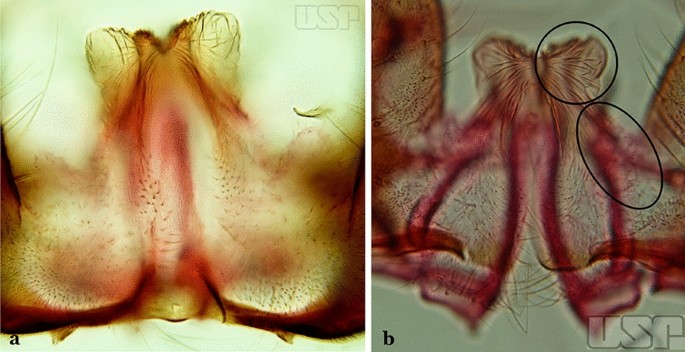
a An. strodei. b An. albertoi
-
38b
Spicules of lateral and dorsal surfaces of apical half of ventral claspette extend to 0.5 of apical portion (Fig. 45b); basoventral lobe of ventral claspette with long and well-developed spicules, spicules denser and longer than on median portion…..An. albertoi
-
39a (34b)
Aedeagus with subapical leaflets (Fig. 46a)…..40
Fig. 46 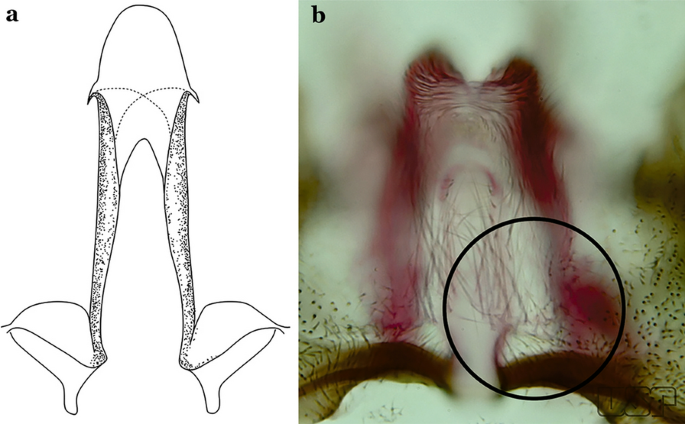
a An. ininii (redrawn after Faran [9]). b An. goeldii Rozeboom & Gabaldon, 1941
-
39b
Aedeagus without subapical leaflets (Fig. 46b)…..42
-
40a (39a)
Ventral claspette clearly conical; basal lobe large and curved in a distal direction, with very long spicules; preapical plate large, strongly sclerotized, half- moon-shaped (Fig. 47a); aedeagus with distal extremity membranous, rounded (Fig. 46a)…..An. ininii
Fig. 47 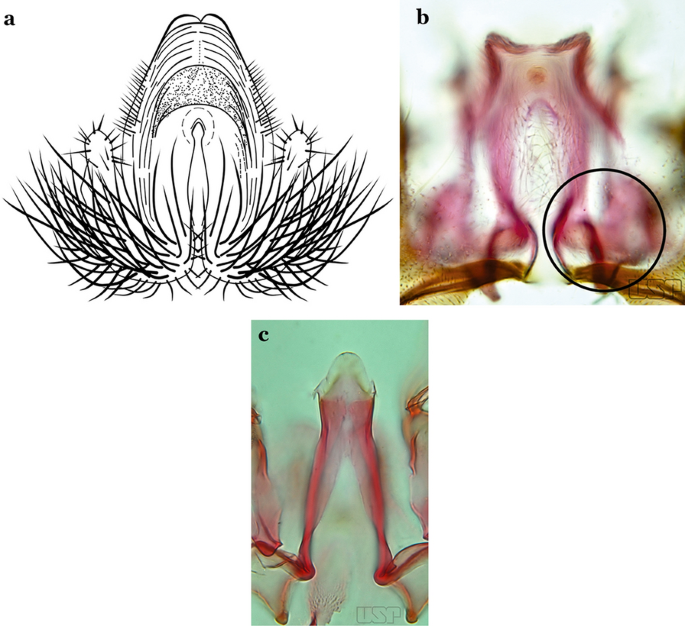
a An. ininii Senevet & Abonnenc, 1938 (redrawn after Faran [9]). b, c An. nuneztovari
-
40b
Ventral claspette rectangular (Fig. 47b); basoventral lobe of ventral claspette rectangular, small, straight, with short spicules (Fig. 46b); preapical plate indistinct (Figs. 46b, 47b); aedeagus with apical extremity membranous, triangular, frequently with small subapical leaflets, more easily visible when aedeagus is separated from other structures of genitalia (Figs. 47c)…..41
-
41a (40b)
Apex of aedeagus more or less triangular (Fig. 47c); basomedian portion of ventral claspette with short sparse spicules (Fig. 47b)…..An. nuneztovari
-
41b
Apex of aedeagus more or less quadrangular (Fig. 48); basomedian portion of ventral claspette with moderately long and dense spicules (Fig. 46b)….. An. goeldii
Fig. 48 
An. goeldii
-
42a (39b)
Ventral claspette with a dense array of spicules on basomedian margin of basoventral lobe; preapical plate small and strongly sclerotized (Fig. 49a)….. An. rangeli
Fig. 49 
a An. rangeli Gabaldon, Cova-Garcia & Lopez, 1940. b An. dunhami Causey, 1945
-
42b
Ventral claspette without a dense array of spicules on basoventral lobe; preapical plate not as above (Fig. 49b)…..43
-
43a (42b)
Ventral claspette with apex broad, quadrangular in outline, forming an angle of about 90° with lateral margin (Fig. 50a)…..An. dunhami & An. trinkae
Fig. 50 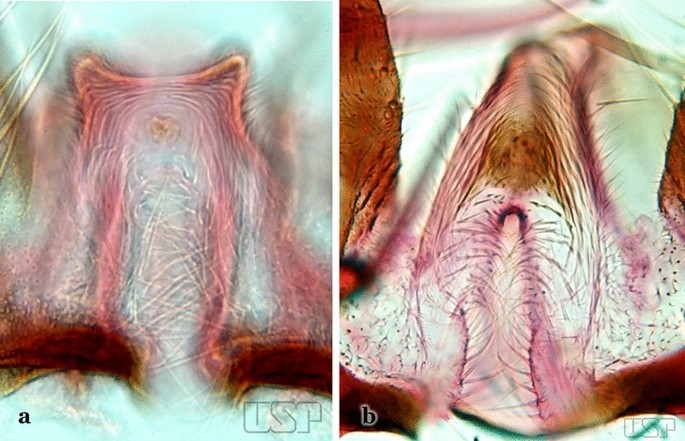
a An. dunhami. b An. galvaoi Causey, Deane & Deane, 1943
-
43b
Ventral claspette with apex narrow, trapezoidal in outline, gradually continuous with lateral margin (Fig. 50b)…..44
-
44a (43b)
Aedeagus with apical portion very short and truncate (Fig. 51a)…..An. evansae
Fig. 51 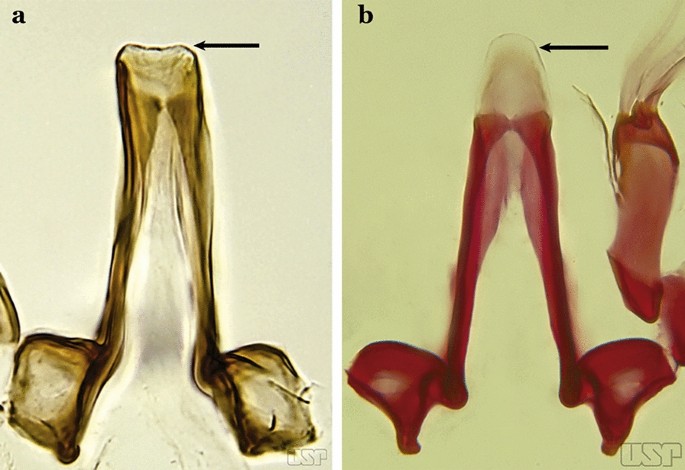
a An. evansae (Brèthes, 1926). b An. galvaoi
-
44b
Aedeagus with apical portion elongate and rounded (Fig. 51b)…..45
-
45a (44b)
Ventral claspette with basoventral lobe rounded, with sparse, moderately long spicules on basal margin; spicules of ventral claspette not extending to apex; preapical plate rounded, moderately sclerotized (Fig. 52a)…..An. aquasalis
Fig. 52 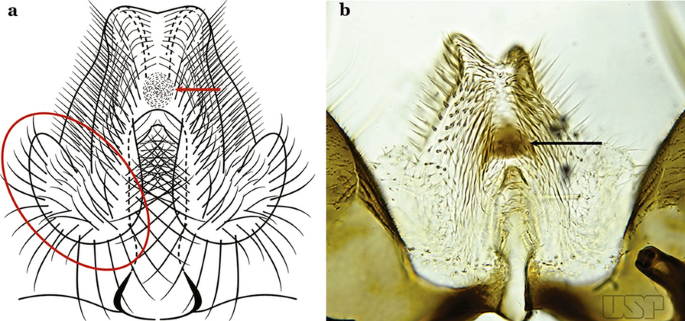
a An. aquasalis Curry, 1932 (redrawn after Faran [9]). b An. konderi Galvão & Damasceno, 1942
-
45b
Ventral claspette with basoventral lobe rounded, with numerous long spicules on basal margin; spicules of ventral claspette extending to apex; preapical plate half- moon-shaped, strongly sclerotized (Fig. 52b)…..46
-
46a (45b)
Ventral claspette with long and numerous spicules on basoventral margin of basal lobe, spicule length up to nearly 3 times width of aedeagus (Fig. 53a); preapical plate large, sclerotized (Fig. 53b)….. An. galvaoi
Fig. 53 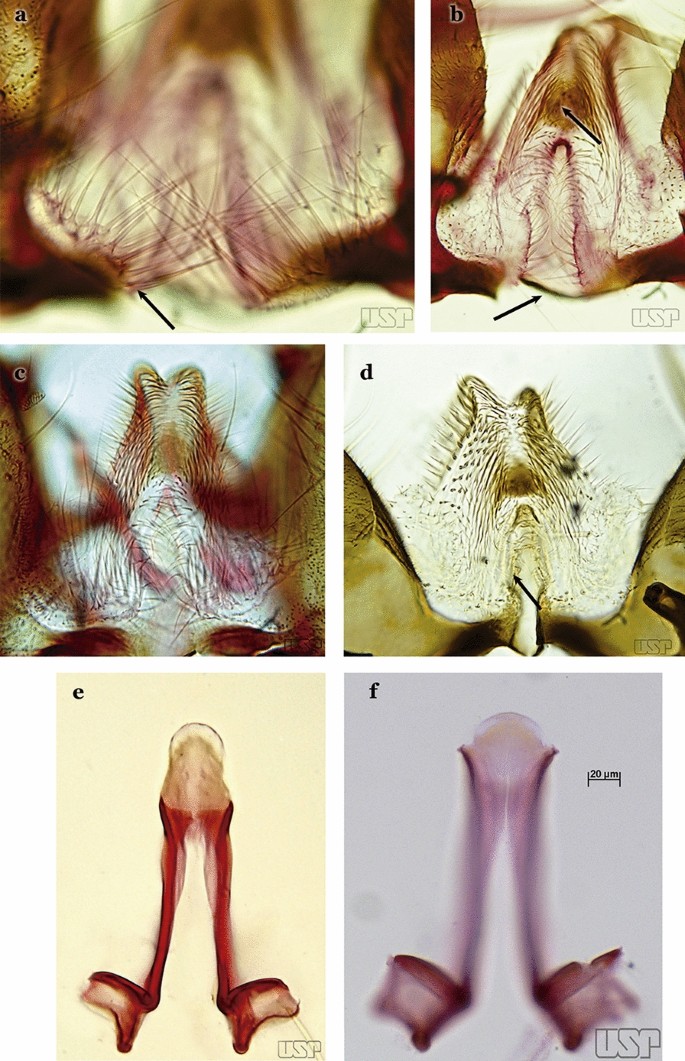
a, b An. galvaoi. c, e An. oswaldoi (Peryassú, 1922). d, f An. konderi
-
46b
Ventral claspette with shorter spicules on basal margin of basoventral lobe, spicule length about 2 times width of aedeagus (Figs. 53c, d); preapical plate large, less sclerotized than described above (Figs. 53e, f)…..47
-
47a (46b)
Apex of aedeagus longer than wide, without subapical points (“leaflets”) (Fig. 53e)…..An. oswaldoi (s.l.)
-
47b
Apex of aedeagus wider than long, with small subapical acute lateral points (“leaflets”) (Fig. 53f)…..An. konderi (s.l.)
-
48a (7b)
Aedeagus with 2 pairs of apical leaflets, but only 1 obvious pair (Fig. 54a)…..An. squamifemur
Fig. 54 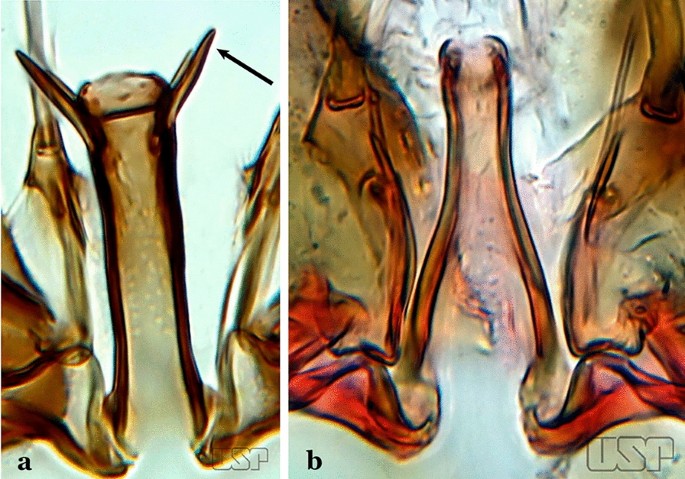
a An. squamifemur Antunes, 1937. b An. pseudotibiamaculatus Galvão & Barretto, 1941
-
48b
Aedeagus without apical leaflets (Fig. 54b)…..49
-
49a (48b)
Apical seta of dorsal claspette expanded, spoon-like, with apex rounded (Fig. 55a)…..50
Fig. 55 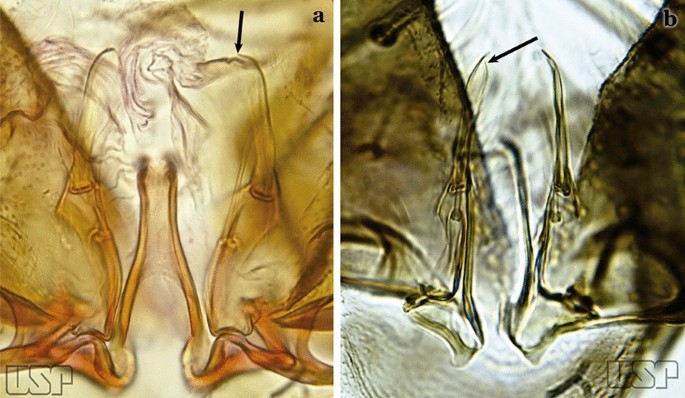
a An. pseudotibiamaculatus. b An. gilesi (Neiva, 1908)
-
49b
Apical seta of dorsal claspette expanded, with apex narrow or lanceolate (Fig. 55b) ………..51
-
50a (49a)
Aedeagus short, curved and conical, well sclerotized, with apex not expanded (Fig. 56a)…..An. pseudotibiamaculatus
Fig. 56 
a An. pseudotibiamaculatus. b An. gomezdelatorrei Levi-Castillo, 1955
-
50b
Aedeagus elongated and tubular, with apex expanded and rounded (Fig. 56b)…..An. gomezdelatorrei
-
51a (49b)
Aedeagus hyaline, with apical extremity expanded, semicircular in outline (Fig. 57a)…..An. vargasi
Fig. 57 
a An. vargasi Gabaldon, Cova-Garcia & Lopez, 1941 (redrawn after Levi-Castillo [15]). b An. gilesi (Neiva, 1908)
-
51b
Aedeagus not hyaline, with apical extremity not expanded (Fig. 57b)…..52
-
52a (51b)
Aedeagus with apical extremity quadrangular in outline (Fig. 58a); strongest seta of dorsal claspette single, not branched (Fig. 58b)….. An. gilesi
Fig. 58 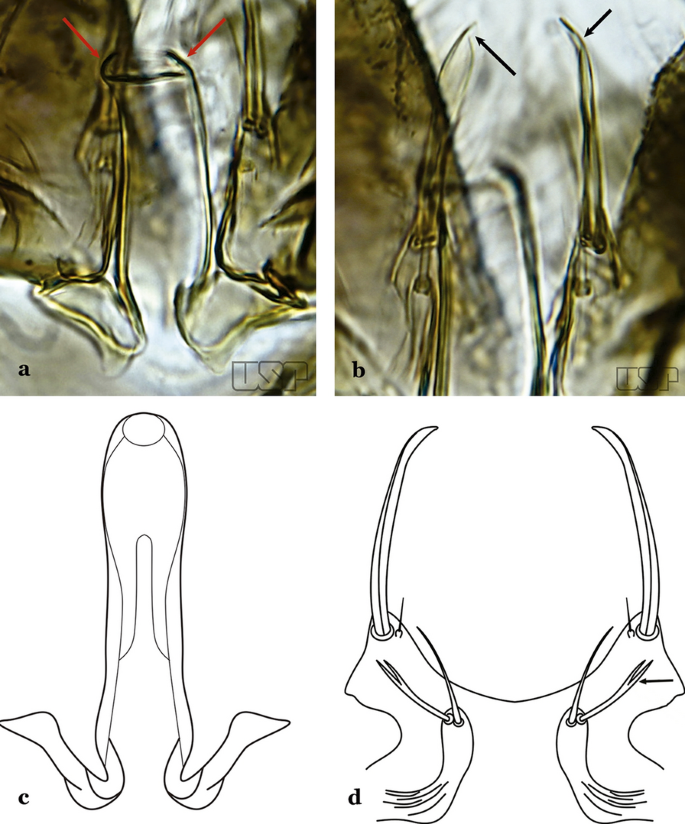
a, b An. gilesi. c, d An. oiketorakras Osorno-Mesa, 1947 redrawn after Lane [16]
-
52b
Aedeagus with apical extremity triangular or trapezoidal in outline (Fig. 58c); strongest seta of dorsal claspette forked (Fig. 58d)…..An. oiketorakras
-
53a (6b)
Aedeagus without apical leaflets, quadrangular in outline, with 2 lateral rod-like sclerotizations connected basally by a hyaline bridge and basally with 2 lateral hyaline digitiform expansions (Fig. 59a)….. An. tibiamaculatus
Fig. 59 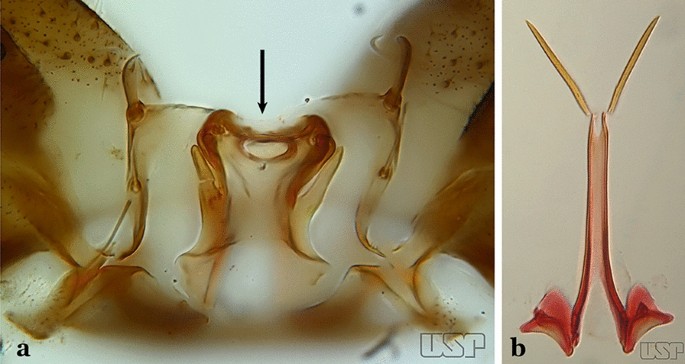
a An. tibiamaculatus Neiva, 1906. b An. costai
-
53b
Aedeagus with apical leaflets of variable appearance (Fig. 59b)…..54
-
54a (53b)
Lobes of tergum IX large, nearly parallel, except apically, approximated medially, space between them narrow (Fig. 60a)…..55
Fig. 60 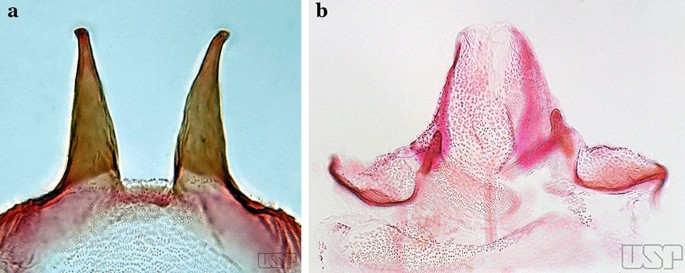
a An. mediopunctatus (Lutz, 1903). b An. eiseni Coquillett, 1902
-
54b
Lobes of tergum IX small, widely separated, space between them large (Fig. 60b)…..57
-
55a (54a)
Ventral claspette nearly as long as wide, with a rounded apex in lateral view, without apicodorsal projection, dorsomedial surface with strong and numerous long spicules (Fig. 61a); lobes of tergum IX with a broad base, narrowing toward apex, lobes moderately divergent (Fig. 61b)…..An. mediopunctatus
Fig. 61 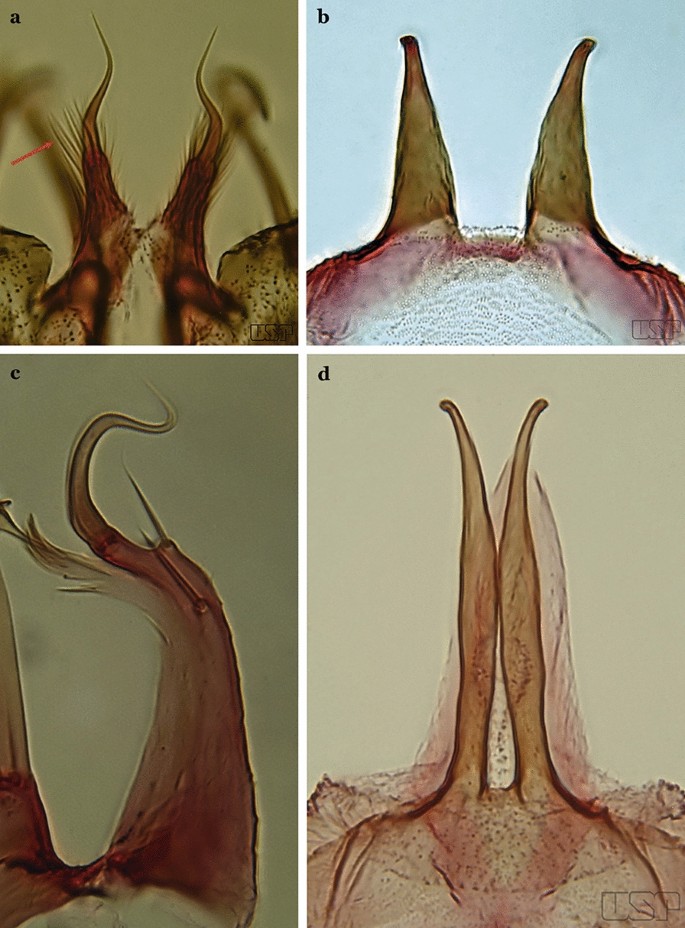
a, b An. mediopunctatus. c An. costai. d An. forattinii Wilkerson & Sallum, 1999
-
55b
Ventral claspette columnar, longer than wide, apex with apicodorsal projections directed posteriorly, dorsomedial surface with small sparse spicules (Fig. 61c); lobes of tergum IX very close together, parallel, divergent in apical 0.3 (Fig. 61d)…..56
-
56a (55b)
Dorsal claspette ovoid, almost circular (Fig. 62a); area between dorsal and ventral claspette U-shaped (Fig. 61d)…..An. costai
Fig. 62 
a An. costai. b An. forattinii redrawn after Wilkerson & Sallum [17]
-
56b
Dorsal claspette nearly triangular, never ovoid; area between dorsal and ventral claspette V-shaped (Fig. 62b)…..An. forattinii
-
57a (54b)
Aedeagus with a single pair of leaflets, with smooth or serrate margins (Fig. 63a)…..58
Fig. 63 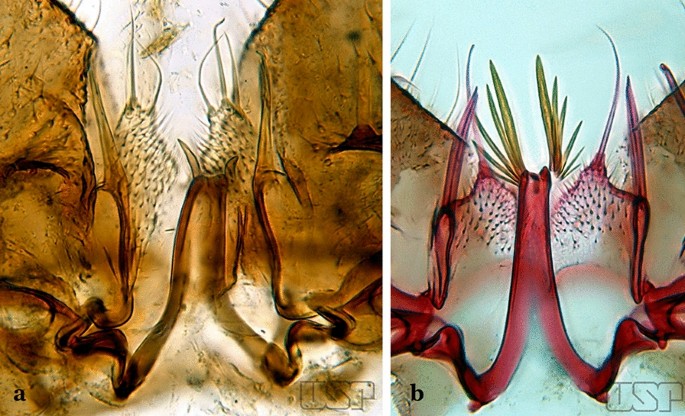
a An. mattogrossensis Lutz & Neiva, 191163. b An. peryassui
-
57b
Aedeagus with more than one pair of leaflets, with smooth or serrate margins (Fig. 63b)…..60
-
58a (57a)
Aedeagus with long leaflets, 0.50 to 0.75 length of aedeagus; ventral claspette spiculose, with 1 stout apical seta and 1 or 2 smaller accessory setae (Fig. 64a); dorsal claspette conical, with 2 or 3 apically spatulate, closely approximated, club-like setae (Fig. 64b)…..An. vestitipennis
Fig. 64 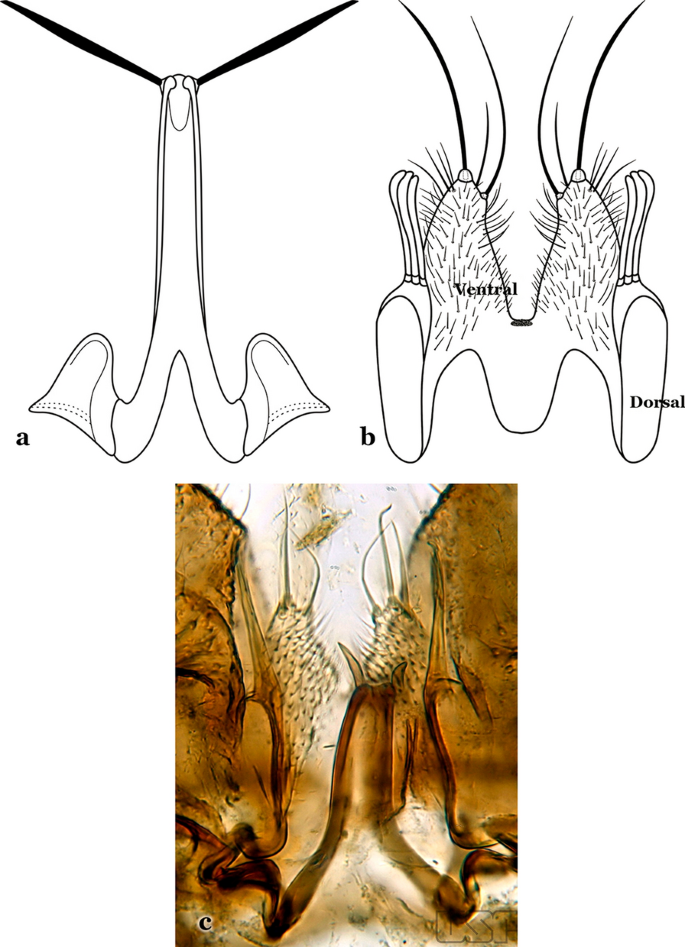
a, b An. vestitipennis Dyar & Knab, 1906 (redrawn after Komp [7]). c An. mattogrossensis Lutz & Neiva, 1911
-
58b
Aedeagus with short leaflets, shorter than 0.5 length of aedeagus (Fig. 64c)…..59
-
59a (58b)
Aedeagus with strongly sclerotized leaflets (Fig. 64c), with smooth margins; dorsal claspette with setae similar in development; ventral claspette as in Fig. 65a…..An. mattogrossensis
Fig. 65 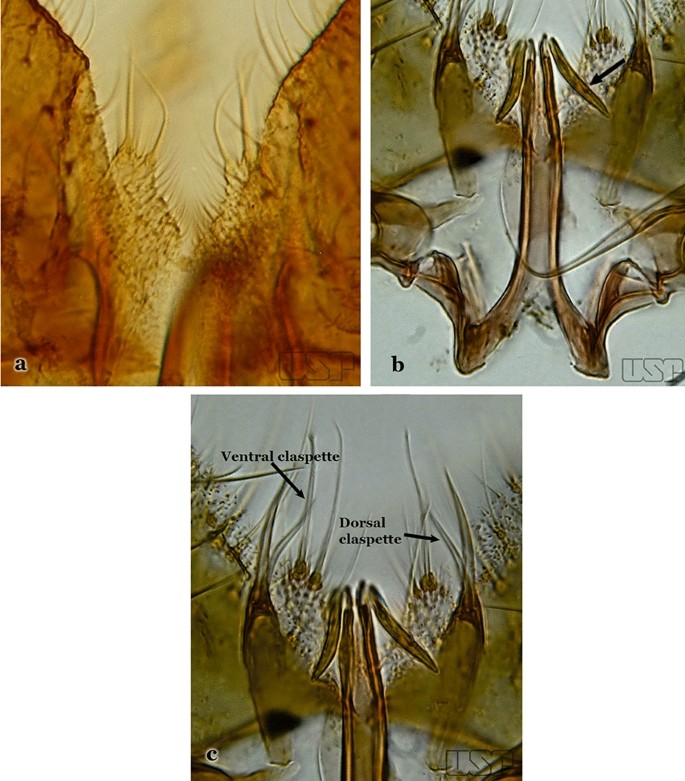
a An. mattogrossensis. b, c An. eiseni geometricus Correa, 1944
-
59b
Aedeagus with leaflets with serrate margins (Fig. 65b); dorsal claspette with 2 or 3 setae, one of which is longer and broader than others (Fig. 65c)…..An. eiseni eiseni & An. eiseni geometricus
-
60a (57b)
Gonocoxite without an internal seta (Fig. 66a)…..61
Fig. 66 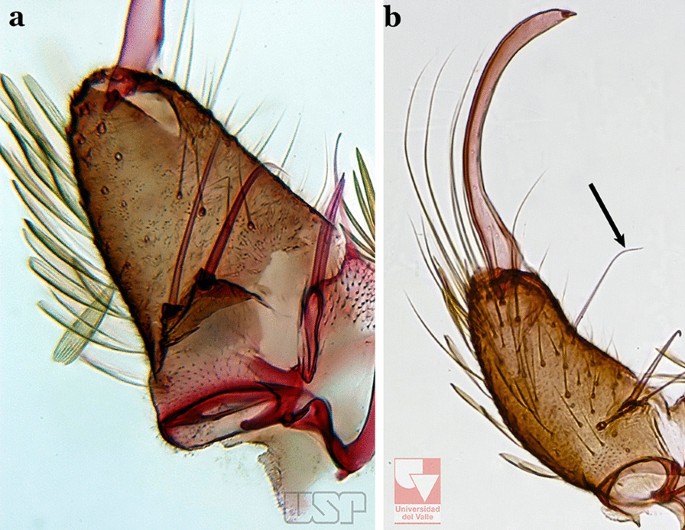
a An. peryassui. b An. calderoni
-
60b
Gonocoxite with an internal seta (Fig. 66b)…..62
-
61a (60a)
Aedeagus elongate, with 4 or more pairs of apical leaflets gradually decreasing in size, terminal leaflet well sclerotized (Fig. 67a); ventral claspette with a long apical seta (Fig. 67b)….. An. peryassui
Fig. 67 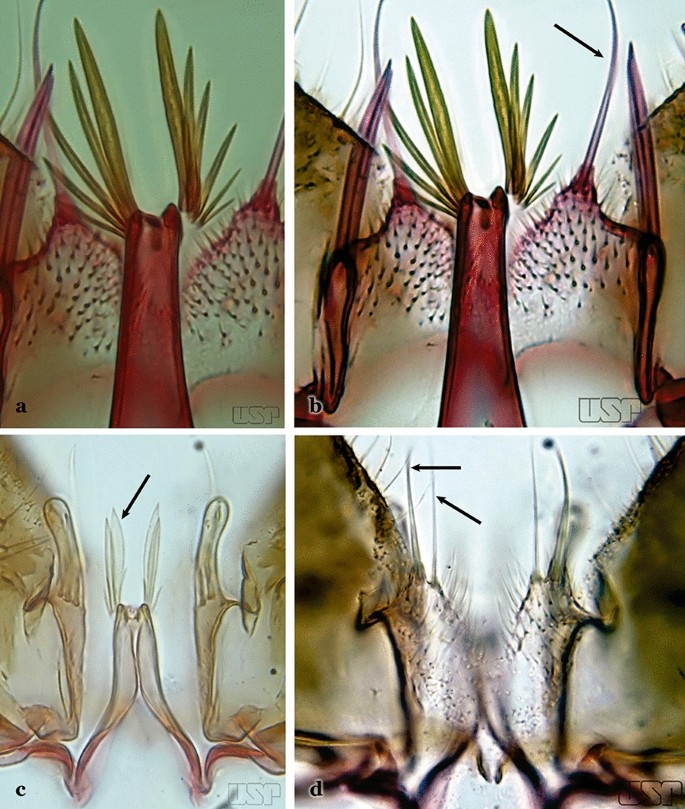
a, b An. peryassui. c, d An. minor da Costa Lima, 1929
-
61b
Aedeagus short, with 2 primary pairs of apical leaflets on each side and 2 or 3 rudimentary ones; ventral claspette with 2 long setae at apex (Figs. 67c, d)….. An. minor
-
62a (60a)
Aedeagus with a pair of broad apical leaflets, remaining leaflets indistinct, variable in number, resembling spicules (Fig. 68a)…..63
Fig. 68 
a An. anchietai Corrêa & Ramalho, 1968. b An. fluminensis Root, 1927
-
62b
Aedeagus with a pair of apical leaflets, narrower than those described above or spiculate, remaining leaflets can vary in number and development, but are always easily visible (Fig. 68b)…..64
-
63a (62a)
Aedeagus with a pair of apical leaflets, size varying from 0.5 to 0.75 length of aedeagus; apical aperture wide and slightly concave (Fig. 69a)…..An. maculipes
Fig. 69 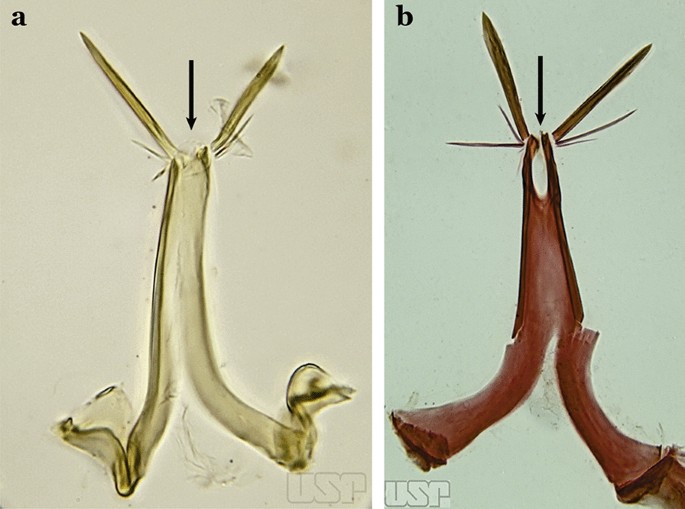
a An. maculipes (Theobald, 1903). b An. anchietai
-
63b
Aedeagus with a pair of long and broad apical leaflets, length < 0.5 length of aedeagus; apical aperture narrow, elongated, deeply U-shape (Fig. 69b)….. An. anchietai
-
64a (62b)
Aedeagus with apical leaflets with smooth margins (Fig. 70a)…..65
Fig. 70 
a An. apicimacula Dyar & Knab, 1906. b An. pseudopunctipennis (both redrawn after Komp [7])
-
64b
Aedeagus with apical leaflets with margins distinctly serrated along their whole length (Fig. 70b)…..69
-
65a (64a)
Aedeagus relatively short, with a pair of long apical leaflets, straight with an oblique apex (Fig. 70a); ventral claspette with long spicules and 3 or 4 setae, one of them longer (Fig. 71a)….. An. apicimacula
Fig. 71 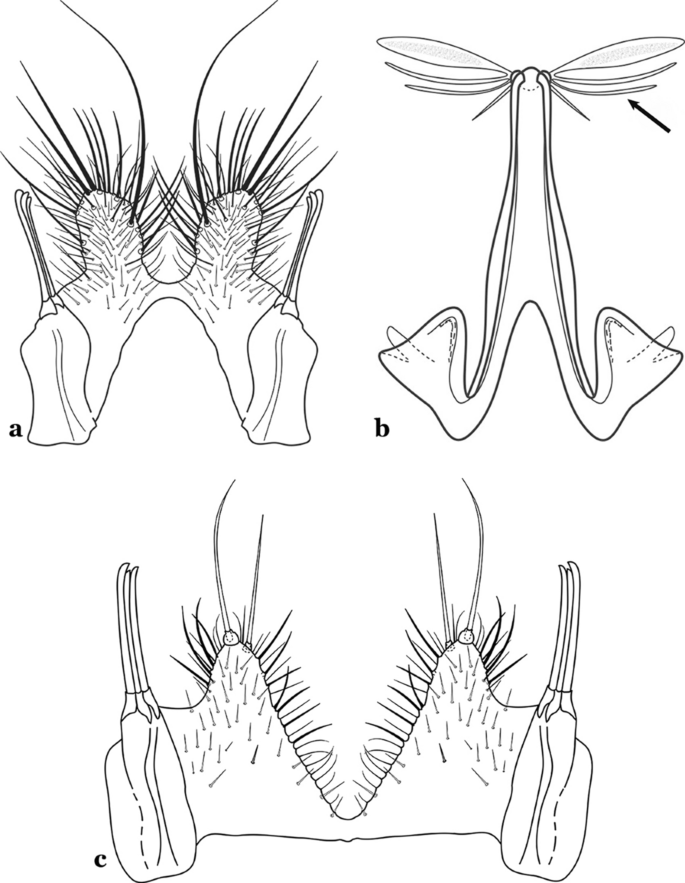
a An. apicimacula. b, c An. punctimacula Dyar & Knab, 1906 (both redrawn after Komp [7])
-
65b
Aedeagus of variable length, with 1 pair of apical leaflets more developed and broader (Fig. 71b); ventral lobe spicules of variable length and number (Fig. 71c)…..66
-
66a (65b)
Aedeagus with apical leaflets more sclerotized medially, margins hyaline (Fig. 72a); ventral claspette with a long slender and curved seta, and another smaller, slender one; dorsal claspette with a long stalk, with 3 strong well- developed setae inserted on apex, setae with curved and pointed apices (Fig. 72b)….. An. calderoni, An. guarao, An. malefactor & An. punctimacula
Fig. 72 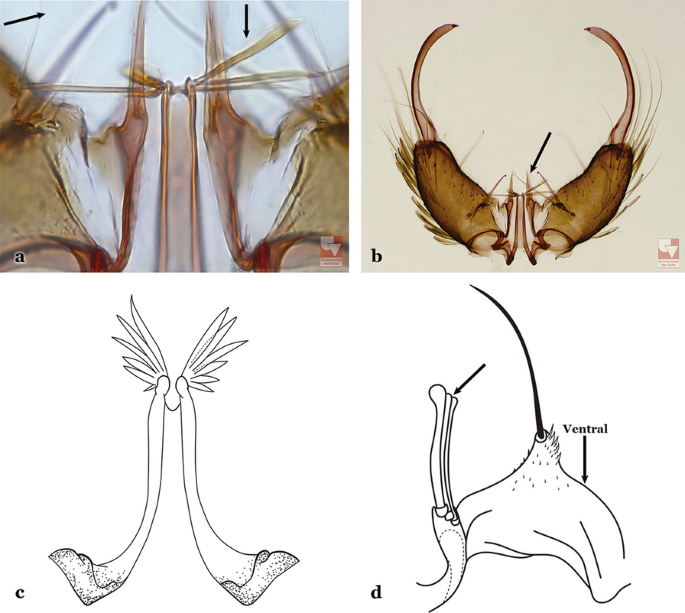
a, b An. calderoni Wilkerson, 1991. c, d An. shannoni Davis, 1931 (redrawn after Wilkerson et al. [18])
-
66b
Aedeagus with a pair of uniformly sclerotized apical leaflets, including margins (Fig. 72c); ventral and dorsal claspette with variable (in number and form) setae inserted on apex (Fig. 72d)…..67
-
67a (66b)
Ventral claspette with a long apical seta (Fig. 72d); aedeagus with 5 to 8 pairs of apical leaflets (Fig. 72c)…..An. shannoni
-
67b
Ventral claspette with more than 1 pair of setae near apex (Fig. 73a); aedeagus with smaller number of apical leaflets (Fig. 73b)…..68
Fig. 73 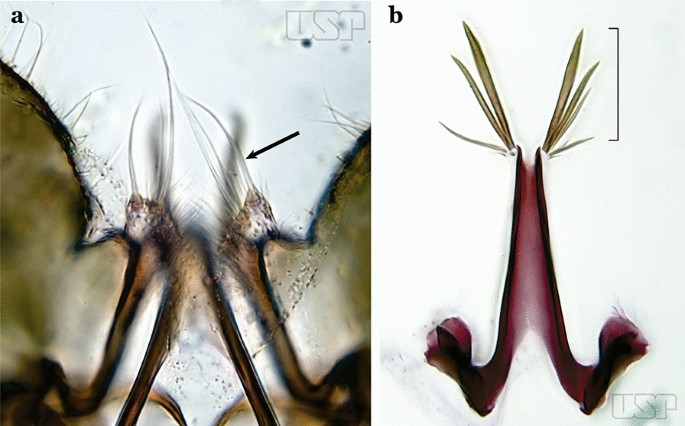
a An. medialis Harbach, 2018 (redrawn after Wilkerson et al. [18]). b An. fluminensis
-
68a (67b)
Aedeagus with at least 3 pairs of small, uniformly sclerotized apical leaflets (Fig. 74a)…..An. medialis
Fig. 74 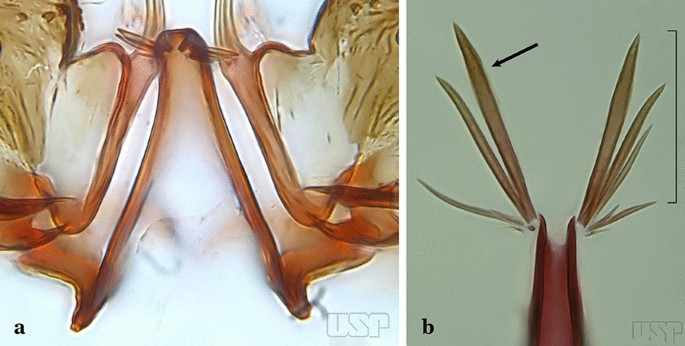
a An. medialis. b An. fluminensis
-
68b
Aedeagus with 5 pairs of long to moderately long apical leaflets with hyaline margins (Fig. 74b)…..An. fluminensis
-
69a (64b)
Aedeagus usually with 4 pairs of leaflets present, apical pair long, sabre- like, serrate or with small spicules at margins, widened beyond middle, tapering to apex, other leaflets slender (Fig. 75a); ventral claspette with 2 long, narrow setae; dorsal claspette with a short stalk, with 3 laminate setae, with apices expanded and slightly curved (Fig. 75b)…..An. neomaculipalpus
-
69b
Aedeagus relatively short, strongly curved dorsally, with more than 1 pair of small, delicate apical leaflets (Fig. 70b); ventral claspette with 2 strong, long, curved setae (Fig. 75c); dorsal claspette with a columnar stalk, with 3 flattened, closely appressed, bladelike setae…..An. pseudopunctipennis




































































Conclusions
Male genitalia possess characters that can be employed for accurate species identification. However, caution is necessary during the entire multi-step preparation procedure, including staining in a solution of acid fuchsine, separation of some parts with extra-fine needles, and repositioning each part on the microscope slide before covering with a coverslip. Any distortion during the dissection and mounting process presents an obstruction for accurate identification. In particular, the ventral claspette must not be distorted, because this will obstruct recognition of its unique features and shape that are essential for identification. For some species, the shape, and anatomical details of the aedeagus must be examined for species identification. For members of the Myzorhynchella Series, the ventral and dorsal claspettes possess multiple characteristics that are here employed for species identification.
Availability of data and materials
Specimens used in the current study are deposited and available in the Coleção Entomológica de Referência, Faculdade de Saúde Pública, Universidade de São Paulo (FSP-USP), São Paulo State, Brazil, the US National Mosquito Collection, Smithsonian Institution, Washington, DC, USA (USNMC), and the Facultad de Ciencias Naturales y Exactas de la Universidad del Valle, Colombia.
References
Sallum MAM, Gonzalez Obando R, Carrejo N, Wilkerson RC. Identification keys to the Anopheles mosquitoes of South America (Diptera: Culicidae). I. Introduction. Parasit Vectors. 2020. https://doi.org/10.1186/s13071-020-04298-6.
Sallum MAM, Gonzalez Obando R, Carrejo N, Wilkerson RC. Identification keys to the Anopheles mosquitoes of South America (Diptera: Culicidae). II. Fourth instar larvae. Parasit Vectors. 2020. https://doi.org/10.1186/s13071-020-04299-5.
Sallum MAM, Gonzalez Obando R, Carrejo N, Wilkerson RC. Identification keys to the Anopheles mosquitoes of South America (Diptera: Culicidae). IV. Adult females. Parasit Vectors. 2020. https://doi.org/10.1186/s13071-020-04301-0.
Harbach RE, Knight KL. Taxonomistsʼ glossary of mosquito anatomy. Marlton: Plexus Publishing, Inc.; 1980.
Harbach RE, Knight KL. Corrections and additions to taxonomistsʼ glossary of mosquito anatomy. Mosq Syst. 1981;1982:201–17.
McAlpine JF, Peterson BV, Shewell GE, Teskey HJ, Vockeroth JR, Wood DM. Manual of Nearctic Diptera, vol. 1. Ottawa: Biosystematics Research Institute; 1981.
Komp WHW. The anopheline mosquitoes of the Caribbean Region. Natl Inst Health Bull. 1942;179:195.
Forattini OP. Culicidologia Médica. Vol. I. Principios gerais, morfologia, glossario taxonômico. São Paulo: Editora da Universidade de São Paulo, EDUSP; 1996.
Faran ME. Mosquito studies (Diptera: Culicidae) XXXIV. A revision of the Albimanus section of the subgenus Nyssorhynchus of Anopheles. Contr Amer Entomol Inst. 1980;15:1–215.
Faran ME, Linthicum KJ. A handbook of the Amazonian species of Anopheles (Nyssorhynchus) (Diptera: Culicidae). Mosq Syst. 1981;13:1–81.
Forattini OP. Culicidologia Medica. Vol. II. Identificacão, Biologia, Epidemiologia. São Paulo: Editora da Universidade de Sao Paulo, EDUSP; 2002.
Komp WHW. The species of the subgenus Kerteszia of Anopheles (Diptera, Culicidae). Ann Entomol Soc Am. 1937;30:492–529.
Zavortink TJ. Mosquito studies (Diptera, Culicidae) XXIX. A review of the subgenus Kerteszia of Anopheles. Contrib Amer Entomol Inst. 1973;9:1–54.
Linthicum KJ. A revision of the Argyritarsis Section of the subgenus Nyssorhynchus of Anopheles (Diptera: Culicidae). Mosq Syst. 1988;20:98–271.
Levi-Castillo R. Un nuevo anofelino de altura del Ecuador: Anopheles gomezdelatorrei n. sp. Revta Ecuator Ent Parasit. 1955;2:509–14.
Lane J. Neotropical Culicidae, vol. I. Sao Paulo: University of Sao Paulo; 1953.
Wilkerson RC, Sallum MAM. Anopheles (Anopheles) forattinii: a new species in the Series Arribalzagia (Diptera: Culicidae). J Med Entomol. 1999;36:345–54.
Wilkerson RC, Sallum MAM, Forattini OP. Redescription of Anopheles (Anopheles) shannoni Davis; a member of the Arribalzagia Series from the Amazon Basin (Diptera: Culicidae). Proc Entomol Soc Wash. 1997;99:461–71.
Acknowledgements
This work would not have been possible without the examination and permission for photographs of specimens from one of the major collections of mosquitoes in South America, repository of valuable reference specimens of multiple species of Anopheles of the Neotropical Region, including primary and secondary type specimens, at the Coleção Entomológica de Referência da Faculdade de Saúde Pública, Universidade de São Paulo, Brazil (FSP-USP). We also thank the Museum of Entomology at the Universidad del Valle, Colombia, for allowing the use of photographic resources and preparing digital images of various parts of its entomological collection. MAMS extends her thanks to the Saúde Pública, Universidade de São Paulo, for their continued support for research projects and the logistics facility for the preparation and maintenance of thousands of specimens of the mosquito collection; and the Fundação de Amparo à Pesquisa do Estado São Paulo for continuous financial support that allowed the execution of hundreds of field sampling efforts for research in the systematics and ecology of mosquitoes (FAPESP Grants #2014/26229-7; #2011/20397-7; #2005/53973-0; CNPq # 301877/2016 to MAMS). RGO and NSC give special thanks to the Facultad de Ciencias Naturales y Exactas de la Universidad del Valle, Colombia, for continuous support and the logistics facility. Project Amazon Malaria Initiative (AMI) - Amazon Network for the Surveillance of Antimalarial Drug Resistance (RAVREDA) provided partial financial support with assistance from USAID and coordination with PAHO/WHO. We are in debt to Yvonne-Marie Linton (Walter Reed Army Institute of Research) and Bruce Harrison (in memoriam) for their thoughtful review of the first version of the identification keys for females, males, and larvae, Caio Cesar Moreira, Faculdade de Saúde Pública, Universidade de São Paulo, for final editing of all illustrations, Ralph E. Harbach (Natural History Museum, London, UK) for his thoughtful revision and valuable contribution for the manuscript, and Aneta Kostadinova for her suggestions, corrections, editorial edits that greatly improved the Part III. The activities undertaken at WRBU were performed in part under a Memorandum of Understanding between the Walter Reed Army Institute of Research (WRAIR) and the Smithsonian Institution, with institutional support provided by both organizations. The views expressed in this article are those of the authors and do not necessarily reflect the official policy or position of the U.S. Army, the Department of Defense, or the U.S. Government.
Funding
This study was funded by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) grant no. 2014/26229-7, CNPq grant no. 301877/2016-5 to MAMS; the Armed Forces Health Surveillance Board – Global Emerging Infectious Disease Surveillance (AFHSB-GEIS) [P0116_19_WR_05 and P0140_20_WR_05].
Author information
Authors and Affiliations
Contributions
MAMS and RCW conceived the study. MAMS, RGO and RCW constructed the identification key. MAMS, RGO and NC prepared all illustrations. MAMS, RCW and RGO wrote the manuscript. All authors revised successive drafts of the key. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Sallum, M.A.M., Obando, R.G., Carrejo, N. et al. Identification key to the Anopheles mosquitoes of South America (Diptera: Culicidae). III. Male genitalia. Parasites Vectors 13, 542 (2020). https://doi.org/10.1186/s13071-020-04300-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-020-04300-1